Background: Pathogenic factors driving obesity-induced fatty liver disease (NAFLD) and insulin resistance are not fully understood. While the mechanisms are unclear, we recognize that high fat diets contribute to obesity, insulin resistance, deleterious ROS production, and fibrogenesis. There is evidence that high fat diets can cause an increased risk of diabetes and consequent cardiovascular disease. The chronic overnutrition from high fat diets is increasingly seen as a risk factor for cardio-metabolic diseases. Previously, we established that pharmacological and herbal medication increase the antioxidant and anti-obesity gene heme oxygenase -1 (HO-1), which reduces inflammatory adipokines and hepatosteatosis. Objective: We examined whether targeting adipocytes to overexpress HO-1 would ameliorate the negative metabolic effects of a prolonged high fat diet (HFD) in mice in hepatocyes as well as adipocytes, to reverse NAFLD and prevent subsequent cardiovascular complications. Methods: Mice were administered lentiviral adiponectin-HO-1 after 20 weeks of a HFD and continued on this diet for 8 more weeks, for a total of 28 weeks of HFD. Mice were treated with HO-1 or a sham virus. Mice were euthanized at 28 weeks. Results: HFD induced adipocyte hypertrophy, increased levels of inflammatory adipokines, and the presence of hepatic lipid droplets and steatosis. The treated mice had less liver lipid droplet accumulation, with normal glucose tolerance and vascular function. These beneficial effects of adipocyte-targeted expression of HO-1 correlated with a reduction in inflammatory adipokines and increased insulin receptor phosphorylation. We have demonstrated that selective increased HO-1 expression in adipose tissue via viral transfection, improved mitochondrial function in both adipose tissue and hepatic tissue in a murine model of high fat diet-induced obesity and NAFLD. Conclusion: This represents a unique therapeutic strategy for treating obesity-associated hepatic steatosis and metabolic syndrome from chronic overnutrition and downstream cardiovascular complications.Background: Pathogenic factors driving obesity-induced fatty liver disease (NAFLD) and insulin resistance are not fully understood. While the mechanisms are unclear, we recognize that high fat diets contribute to obesity, insulin resistance, deleterious ROS production, and fibrogenesis. There is evidence that high fat diets can cause an increased risk of diabetes and consequent cardiovascular disease. The chronic overnutrition from high fat diets is increasingly seen as a risk factor for cardio-metabolic diseases. Previously, we established that pharmacological and herbal medication increase the antioxidant and anti-obesity gene heme oxygenase -1 (HO-1), which reduces inflammatory adipokines and hepatosteatosis. Objective: We examined whether targeting adipocytes to overexpress HO-1 would ameliorate the negative metabolic effects of a prolonged high fat diet (HFD) in mice in hepatocyes as well as adipocytes, to reverse NAFLD and prevent subsequent cardiovascular complications. Methods: Mice were administered lentiviral adiponectin-HO-1 after 20 weeks of a HFD and continued on this diet for 8 more weeks, for a total of 28 weeks of HFD. Mice were treated with HO-1 or a sham virus. Mice were euthanized at 28 weeks. Results: HFD induced adipocyte hypertrophy, increased levels of inflammatory adipokines, and the presence of hepatic lipid droplets and steatosis. The treated mice had less liver lipid droplet accumulation, with normal glucose tolerance and vascular function. These beneficial effects of adipocyte-targeted expression of HO-1 correlated with a reduction in inflammatory adipokines and increased insulin receptor phosphorylation. We have demonstrated that selective increased HO-1 expression in adipose tissue via viral transfection, improved mitochondrial function in both adipose tissue and hepatic tissue in a murine model of high fat diet-induced obesity and NAFLD. Conclusion: This represents a unique therapeutic strategy for treating obesity-associated hepatic steatosis and metabolic syndrome from chronic overnutrition and downstream cardiovascular complications.
*Corresponding Author:
Stephen J. Peterson, MD, MACP, FAHA.
Department of Medicine, Weill Cornell Medicine, NY, USA, Tel: 718-780-5246; E-mail: stp9039@med.cornell.edu
ABSTRACT
Background: Pathogenic factors driving obesity-induced fatty liver disease (NAFLD) and insulin resistance are not fully understood. While the mechanisms are unclear, we recognize that high fat diets contribute to obesity, insulin resistance, deleterious ROS production, and fibrogenesis. There is evidence that high fat diets can cause an increased risk of diabetes and consequent cardiovascular disease. The chronic overnutrition from high fat diets is increasingly seen as a risk factor for cardio-metabolic diseases. Previously, we established that pharmacological and herbal medication increase the antioxidant and anti-obesity gene heme oxygenase -1 (HO-1), which reduces inflammatory adipokines and hepatosteatosis. Objective: We examined whether targeting adipocytes to overexpress HO-1 would ameliorate the negative metabolic effects of a prolonged high fat diet (HFD) in mice in hepatocyes as well as adipocytes, to reverse NAFLD and prevent subsequent cardiovascular complications. Methods: Mice were administered lentiviral adiponectin-HO-1 after 20 weeks of a HFD and continued on this diet for 8 more weeks, for a total of 28 weeks of HFD. Mice were treated with HO-1 or a sham virus. Mice were euthanized at 28 weeks. Results: HFD induced adipocyte hypertrophy, increased levels of inflammatory adipokines, and the presence of hepatic lipid droplets and steatosis. The treated mice had less liver lipid droplet accumulation, with normal glucose tolerance and vascular function. These beneficial effects of adipocyte-targeted expression of HO-1 correlated with a reduction in inflammatory adipokines and increased insulin receptor phosphorylation. We have demonstrated that selective increased HO-1 expression in adipose tissue via viral transfection, improved mitochondrial function in both adipose tissue and hepatic tissue in a murine model of high fat diet-induced obesity and NAFLD. Conclusion: This represents a unique therapeutic strategy for treating obesity-associated hepatic steatosis and metabolic syndrome from chronic overnutrition and downstream cardiovascular complications.
Keywords: Heme-oxygenase; inflammation; antioxidants; mitochondrial; insulin resistance’ NAFLD; NASH; cardiometabolic diseases.
ABBREVIATIONS
BP - blood pressure
BVR - biliverdin
CEC - circulating endothelial cell
CO - carbon monoxide
FGF21- Fibroblast growth factor 21
GFR - glomerular filtration rate
GLUT 4 – glucose transporter 4
HO-1 - heme oxygenase isozyme 1
IL – interleukin
IL-10- Interleukin 10
IL-6 Interleukin 6
MCP-1 - monocyte chemoattractant protein-1
MFN1- Mitofusin-1
MFN2- Mitofusin-2
A.K.T. - Protein Kinase (Activator)
Akt - serine/threonine protein kinase
MMP9- Matrix Metallopeptidase 9
NADPH - nicotinamide adenine dinucleotide phosphate (reduced form)
NASH- Non-alcoholic steatohepatitis
NAFLD-Non-alcoholic fatty liver disease
CCN3/NOV- Nephroblastoma overexpressed
PGC1a- Peroxisome proliferator-activated receptor gamma coactivator 1 a
ROS - reactive oxygen species
UCP1- Uncoupling protein 1
WAT-white adipose tissue
INTRODUCTION
The obesity prevalence in the United States in 2017– 2018 reached 42.4% among the adult population [1]. Health risks and complications associated with obesity include hypertension, cardiovascular disease, metabolic syndrome, dyslipidemia, insulin resistance, type 2 diabetes (T2D), and non-alcoholic fatty liver disease (NAFLD) [2-4]. NAFLD is characterized by the accumulation of lipid droplets in the liver and may progress to non-alcoholic steatohepatitis (NASH), which is identified by hepatic inflammation and possible progression to fibrosis, and eventually hepatic cirrhosis and liver failure [5]. High-fat diet and sedentary lifestyle, common in developed countries, have increased NAFLD incidence [6] affecting approximately one-third of the US population, of which 2-5% progress to NASH [7]. High fat diets can contribute to obesity, insulin resistance, deleterious ROS production, and fibrogenesis. There is evidence showing that the overnutrition derived from high fat diets contributes to increased risk of diabetes and cardiovascular disease, and that high-fat diets and high-carbohydrate diets are equal harmful in the progression of liver injury in a murine model of NAFLD [8, 9]. High fat diet regimens have been used to study obesity in rodents, although this is an imperfect model of human obesity [10]. High-fat diet and overnutrition reduce HO-1 levels, increase insulin resistance and impair mitochondrial function in multiple organs, including the liver [11, 12]. Chronic overnutrition also increases serum LDL and Angiotensin II [ANG II] levels, which exacerbate mitochondrial dysfunction and leptin resistance [13].
Overall, the effectiveness of pharmacological intervention for patients with NAFLD remains inadequate, and a significant number of patients progress to NASH [14].
NAFLD progression to NASH is strongly associated with obesity; excess visceral fat disrupts blood glucose homeostasis, leading to insulin resistance and the subsequent appearance of T2D [15], free fatty acid accumulation, chronic inflammation, dyslipidemia, and vascular dysfunction [16-18]. The exact mechanism of hepatic lipid accumulation evolving into NASH has not been fully elucidated, mainly due to limitations in animal models that could consistently replicate the features of human NASH [19, 20]. Genetics and inflammation play an essential role in developing NASH and cardiovascular diseases [20, 21]. Moreover, genetic polymorphisms in HO-1 and HO-1 gene suppression are known to complicate the course of obesity and its evolution to NAFLD/NASH in certain patients [22, 23].
It has been demonstrated that exposing adipocytes to pro-oxidants and reducing HO-1 results in adipocyte hypertrophy and increased production of adipocytokines; the cytokines TNFα, IL-6, MCP- 1, and NOV were all found to be dysregulated in mice and humans [24-28]. Obesity is a chronic inflammatory state that is characterized by immune-mediated inflammation that results in increased reactive oxygen species [ROS] production and suppression of HO-1. [24]. HO-1 deficiency results in increased cellular pro-inflammatory markers, heme, iron deposits, ROS, cellular fibrosis, organ failure, and mortality in human and rodents [29-31]. Oxidative stress is not counteracted by the antioxidant response and the inflamed state of visceral adipocytes causes low levels of adiponectin and insulin resistance [12, 26].
HO-1 degrades heme into biliverdin/bilirubin, carbon monoxide and free iron, which protect cells from the products of inflamed adipocytes [reviewed in [32]]
In the present work, we used an adiponectin promoter-driven lentiviral vector to directly target adipocytes. We hypothesized that adipocyte-targeted expression of HO-1, would improve adipocyte function, reverse hepatic fibrosis, and decrease NASH scores in mice fed a HFD. This study is important because targeting adipocytes with HO-1 would improve both adipocyte function as well as improving hepatic function, which is novel. We propose that adipocyte-targeted expression of HO-1 can represent a potential alternative approach to the treatment of obesity induced NAFLD/NASH, an approach that could last for nine months.
MATERIALS AND METHODS
Experimental animals
All experimental protocols utilizing animals followed the NYMC IACUC institutionally approved guidelines in accordance with the NIH Guide regarding the Care and Use of Laboratory Animals (Protocol #22-2-0415H, 18 April 2018). We used 4-week-old male C57BL/6J mice (Jackson Labs, Bar Harbor, ME). The mice were divided into 3 different treatment groups randomly (N=5 for each group) as follows: 1) Lean, 2) HFD: mice fed an HFD for 20 weeks and 3) HFD: mice fed an HFD for 20 weeks and then injected with adiponectin lentivirus HO-1 (ln-Adipo-HO-1). Mice in group 3 were administered a bolus injection of 80-100 μl Adipo-HO-1 lentivirus (40-70 × 109 TU/ml in saline); and then continued on a HFD for 8 more weeks. The HFD mice (group '2') were similarly injected with sham Ln particles of 1-1.4x109 at 20 weeks and continued on a HFD for 8 more weeks as a control (week 20-28). All mice were euthanized at 28 weeks. (Dharmacon, Lafayette, CO) injected into the retro-orbital vein, and lean mice (group ‘1’) received a mock virus (placebo); all mice in the experimental groups 2 and 3 were kept on HFD for weeks 20-28 while receiving the treatment or sham virus. Control mice (group 1) were fed ad libitum a normal diet containing 11% fat, 62% carbohydrate and 27% protein with a total calorie count of 12.6KJ/g. The remaining animals (groups 2,3) were fed a high-fat diet containing 58% fat (from lard), 25.6% carbohydrate, and 16.4% protein for a total calorie count of 23.4KJ/g. (Bio-SERV, Frenchtown, NJ)
Lentiviral vectors under an adiponectin specific promoter expressing either HO-1 or sham were constructed as described (4) using the LentiMaxTM system (Lentigen, Baltimore, MA and Vector Builder, Shenandoah, TX 77384, USA). Mice were weighed every week and the final body weight was measured at the time of sacrifice. Blood samples were withdrawn and centrifuged for 5 minutes at 12,000g.
Histomorphological analysis
Liver samples were collected from each experimental group and fixed in 4% paraformaldehyde, dehydrated, embedded in paraffin wax, and sectioned (6 μm thick) [33, 34]. The sections were deparaffinized, rehydrated and stained with Masson’s trichrome staining. Stained liver tissue sections were analyzed using an optical microscope (Olympus, Hamburg, Germany) and photomicrographs captured by a digital camera were then morphologically evaluated.
The morphometrical analyses to evaluate NAS pathological factors were performed evaluating 20 randomly chosen liver fields per experimental animal by two independent observers blinded to the experimental group, using computer image analysis software (Image Pro Premier 9.1, Media Cybernetics, USA). In detail, the hepatic steatosis was assessed on stained sections by counting randomly chosen 100 lipid droplets/groups and selecting their diameter (μm) at a final magnification of 400X [34, 35] and liver perivascular fibrosis evaluation, assessed by Masson’s trichrome, normalized to vessel size [36].
Western Blot and qRT-PCR analysis
Adipose and liver tissues were lysed in RIPA lysis buffer supplemented with protease and phosphatase inhibitors. (CompleteTM Mini and PhosSTOPTM, Roche Diagnostics, Indianapolis, IN). Frozen mouse adipose tissue was homogenized in liquid nitrogen and suspended in homogenization buffer (comprising mmol/L:10 phosphate buffer, 250 sucrose,1.0 EDTA, 0.1 PMSF and 0.1%v/v tergitol, pH 7.5). We measured the protein concentration of the supernatant by the Bradford Protein assay (Bio-Rad, Hercules, CA, USA). For Western blot analysis, twenty micrograms of protein extract were separated using SDS-PAGE gels and transferred to a nitrocellulose membrane. After blocking, We incubated the membranes, after blocking, the membranes at 4℃ overnight with the following primary antibodies: HO-1 (Enzo Life Sciences, Farmingdale, NY, USA), MMP9, PRDM16, SIRT1, CREG1, FGF21, MFN2, OPA1, p-AMPK, AMPK, p-ACC, ACC, p-AKT, A.K.T., IR-β, GLUT2 (Cell Signaling Technology, Danvers, MA, USA). We then carried out detection of immunoreactive protein bands as previously described values [4, 37, 38].
Total RNA was obtained from frozen adipose tissue by RNeasy Lipid Tissue (Qiagen), according to the manufacturer's instructions, and reverse transcription was carried out using the High Capacity cDNA Reverse Transcription Kit (Applied BiosystemsTM, Waltham, MA, US). PCR reactions were performed in 10 µL volumes using PowerUp SYBR Green Master Mix (Applied BiosystemsTM) at a concentration of 0.05 µM as described (4, 37, 38).
Statistical analysis
Statistical significance between experimental groups was determined by ANOVA with Tukey-Kramer post-hoc analysis for comparison between multiple groups (GraphPad Prism). The data are presented as means ± SEM, and p<0.05 was considered statistically significant.
RESULTS
Targeting adipocyte specific expression of HO-1 restores hepatocytes
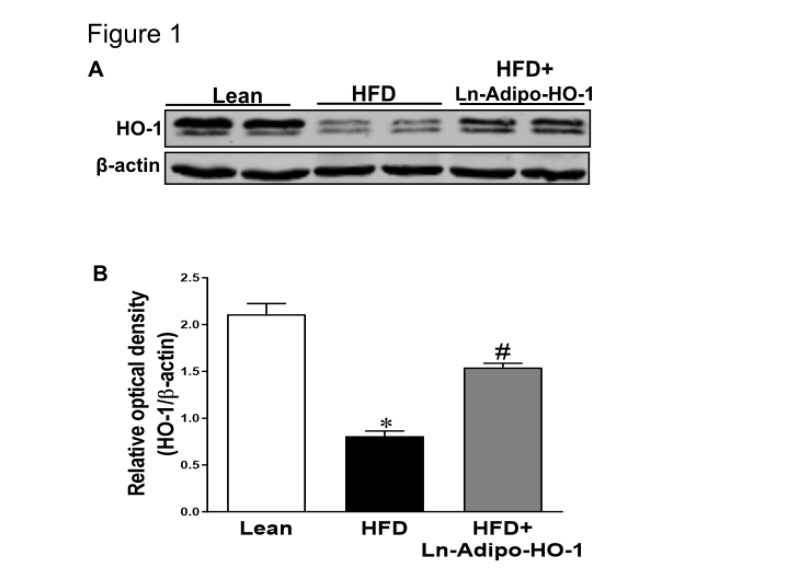
Comparative analysis of HO-1 levels in Lean, HFD, and HFD mice treated with HO-1 indicated HFD significantly (P<0.05) decreased adipocyte HO-1 levels compared to the Lean group. Notably, HO-1 transfection restored (P<0.05) the levels of HO-1 in adipocytes (Figure 1A and B). p<0.05, n=5).
Adipocyte specific HO-1 expression decreases hepatic steatosis and fibrosis
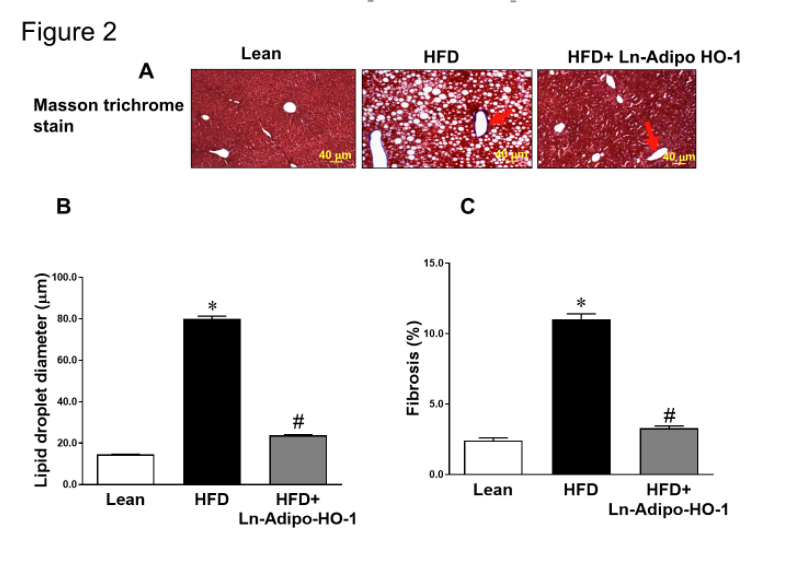
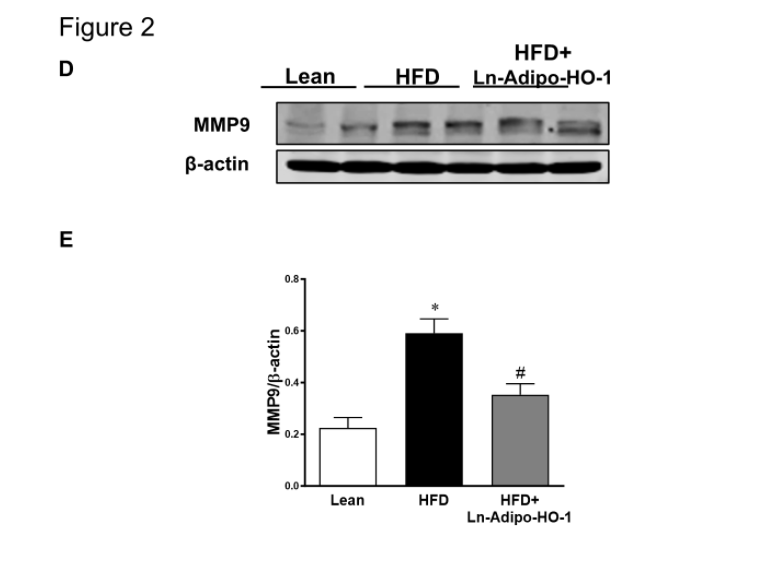
We observed a marked reduction in hepatic lipid droplet diameter and steatosis in HFD fed mice following adipo-HO-1 upregulation (Figure 2A and B). Reduced perivascular fibrosis in the liver was also evident in the HFD+HO-1 group (Figure 2A and C). Furthermore, hepatic levels of MMP9, increased in the HFD group, were attenuated by HO-1 overexpression (Figure 2D-E). This improvement suggests an antifibrotic mechanism following the induction of HO-1.
As expected, livers of lean mice showed no evidence of steatosis, inflammatory foci, or fibrosis, and only rare ballooning, presenting a NAFLD score (NAS) of 1. In contrast, livers of HFD fed mice had a higher NAS value (score of 8). They showed elevated steatosis (49.35% - score 2), moderate lobular inflammatory foci (score 2), significant hepatocyte ballooning (score 2) and perivascular fibrosis (score 2) (Table 1). The enlarged hepatocytes in the HFD group exhibited the hepatocellular ballooning feature. In contrast, control mice presented an average ballooning value of 1.52% ± 0.9 compared to HFD fed mice with a significantly higher value of 27.34% ± 1.12. Adipo-HO-1 expression attenuated NAFLD and showed improvements across all spectra examined, including a reduced NAS value (score 3), low steatosis (score 1), few inflammatory foci (score 1), rare ballooning (score 1), and no fibrosis (score 0) (Table 1). Notably, the hepatocyte ballooning value in HFD+ HO-1 mice was 8.52% ± 1.07. These findings strongly support the notion that upregulation of adipo-HO-1 reversed liver dysfunction, and prevented the development of NAFLD and NASH.
Adipocyte HO-1 expression regulates glucose intolerance and hepatic dysfunction
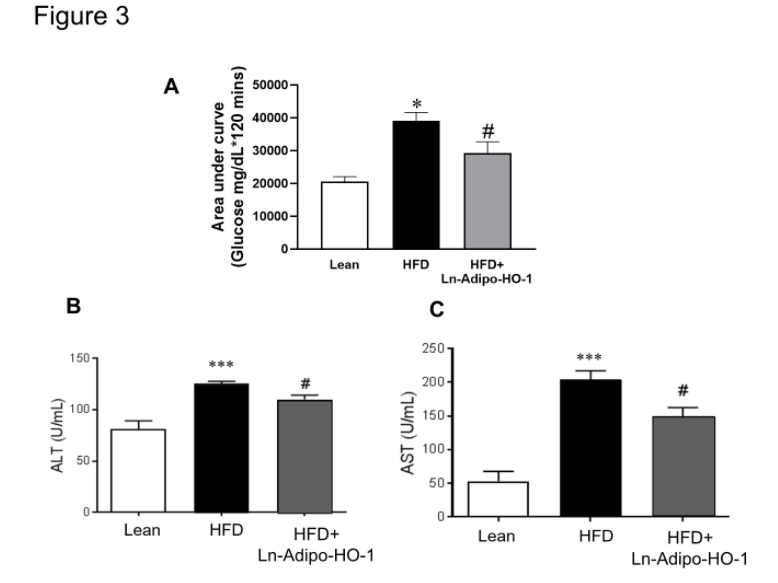
The area under the curve (AUC) glucose indicated that HO-1 overexpression significantly (p<0.05) improved glucose tolerance as compared to the HFD group (Figure 3A). Impairment of liver function in mice fed HFD was clearly shown by increased levels of the serum enzymes AST (p<0.05) and ALT (p<0.05). In agreement with the histological findings, Ln-adipo-HO-1 expression improved liver function, as evidenced by decreased levels of AST and ALT (p<0.05) (Figure 3B and C). n=5
HO-1 upregulation increases genes and proteins related to mitochondrial biogenesis and thermogenesis
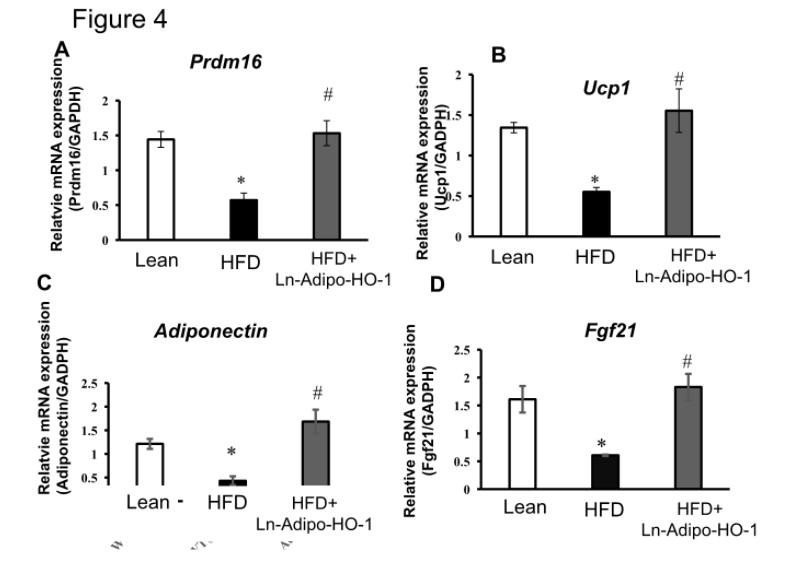
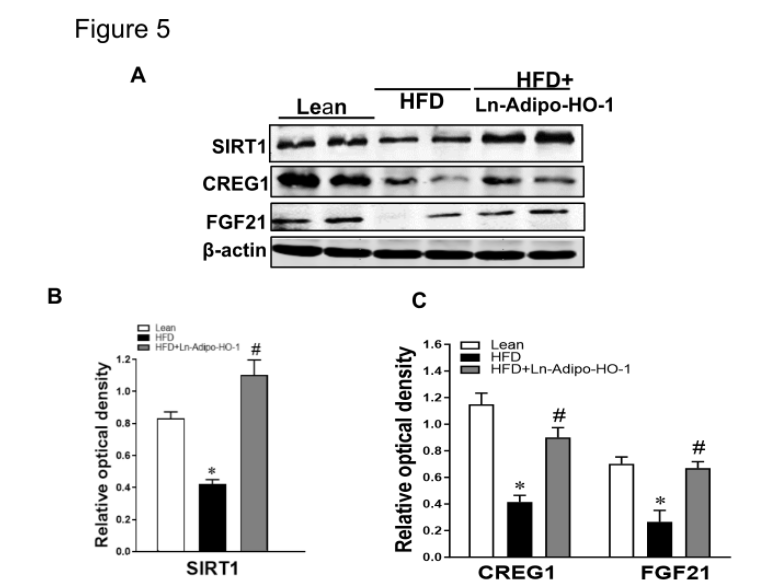
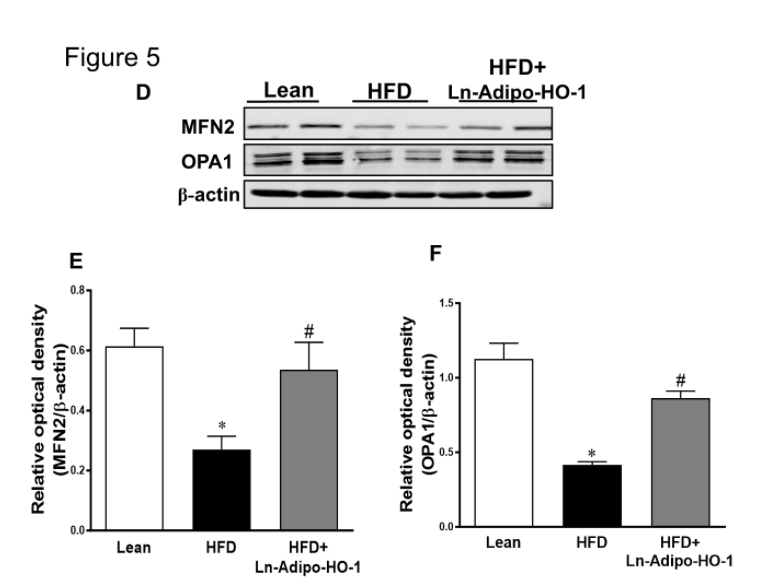
We further measured the expression of genes involved in cellular metabolism and mitochondrial function. As shown in Figure 4A-D, mRNA levels of PRDM16, UCP1, Adiponectin, and FGF21 in adipose tissue of HFD fed mice were decreased (p<0.05) compared with lean mice. This downregulation was prevented in Ln-Adipo-HO-1-HFD mice (p<0.05). Figure 5 also shows that protein levels of SIRT1, CREG1, FGF21, MFN2, and OPA1 were all significantly (p<0.05) increased in the HFD+HO-1 group when compared to the HFD group. These results strongly suggest Ln-adipo-HO-1 expression reduced inflammation and fibrosis by increasing the anti-adiposity genes CREG1, FGF21, thermogenic gene UCP1, and adiponectin, with a subsequent increase in mitochondrial biogenesis and fusion. n=5
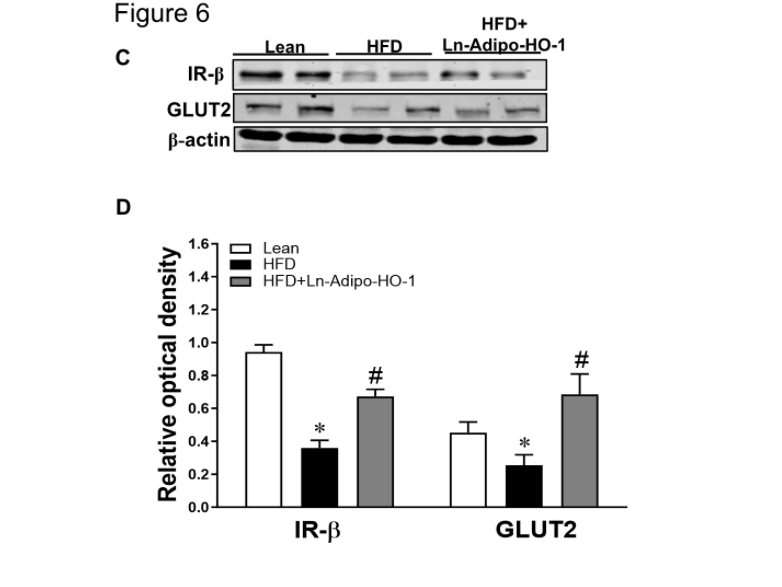
HO-1 overexpression in adipose tissue increases insulin signaling
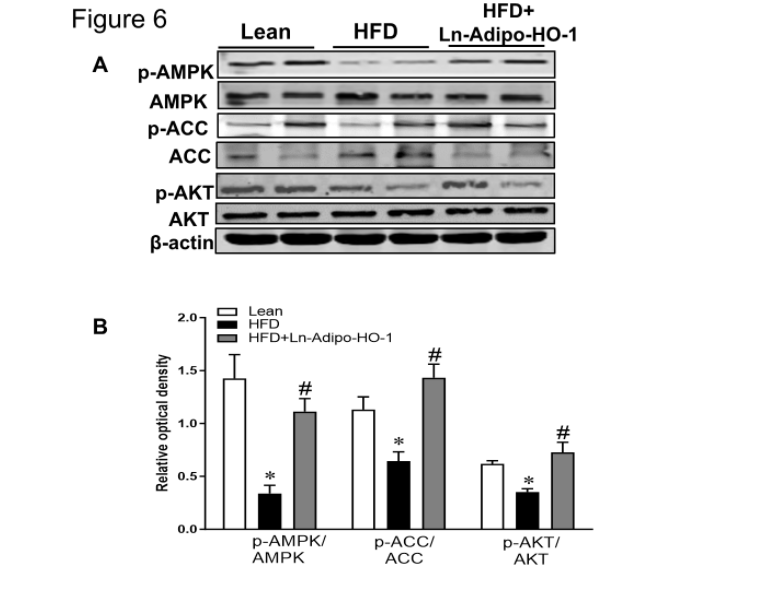
Administration of the Ln-adipo-HO-1 resulted in increases in the upstream signaling of insulin phosphorylation, including AMPK, ACC and AKT Figure 6A and B shows that Ln-adipo-HO-1 expression augmented the phosphorylation status of these proteins, as evidenced by the phosphorylation levels of p-AMPK, p-ACC and p-AKT in liver tissue of mice fed with HFD and treated with Ln-adipo-HO-1, compared to mice fed HFD alone (p<0.05).
An increase in the levels of IR-β and GLUT2 (p<0.05) followed the activation of AMPK and AKT in the HFD Ln-adipo-HO-1 mice, which were downregulated (p<0.05) in the HFD mice (Figure 6C and D). This data suggests that AMPK activation and the subsequent upregulation of the insulin signaling pathway, play a crucial role in mediating the protective effect of HO-1. n=5
DISCUSSION
In the present study, we examined whether adipocyte specific HO-1 overexpression alters liver fat, inflammation and fibrosis. Mice fed a high fat diet were treated with a lentiviral- adiponectin-HO-1 that overexpressed HO-1 in adipose tissue, compared to control. The obesity induced by the high fat diet caused inflammatory changes in both adipocytes and hepatocytes. We have demonstrated the novel finding that selective increased HO-1 expression in adipose tissue via viral transfection, improved mitochondrial function in both adipose tissue and hepatic tissue in a murine model of high fat diet-induced obesity and NAFLD. In addition, we provide evidence demonstrating that adipocyte-specific expression of HO-1 in obese mice displayed pronounced metabolic benefits in both adipocytes and liver, as evidenced by upregulated HO-1 expression in both cell types. This was accompanied by a reduction in hepatic lipid droplets and steatohepatitis and improved glucose tolerance. The remarkable alleviation of the hepatic steatosis did not seem to be associated with changes in food consumption, as all animals consumed the same amount of food (4g/day) regardless of the treatment received. We showed that metabolic improvements were associated with increased levels of UCP1, PRMD16, Sirt1, and CREG1, all with a central role in thermogenesis and mitochondrial signaling. Notably, upregulation of these proteins is required for the increased number and maintenance of brown-like adipocyte stem cells [39, 40]. For the last decade, the browning of white fat has been envisioned as a potential target to develop protection against cardio-metabolic diseases such as obesity [41]. This tissue targeted upregulation of the adipocyte HO-1 signaling pathway also reprogramed the white adipose tissue to a brown-like phenotype and may represent a unique, sustained therapeutic strategy to treat Metabolic Syndrome and NAFLD induced by overnutrition. Pathological numbers of fat white cells are associated with high pro-inflammatory markers and mitochondrial dysfunction [34]. The small adipocytes in brown fat reduce fatty acid accumulation by promoting mitochondrial biogenesis via adaptive mitochondrial thermogenic genes, such as UCP-1 and PRDM-16. UCP-1 uncouples the proton gradient generated by oxidative phosphorylation and promotes energy dissipation in the form of heat, enhancing caloric burning, oxygen consumption, and favoring weight loss [4, 39]. Fibroblast growth factor 21 (FGF21) plays an essential role in regulating the conversion of white adipose tissue (WAT) to brown-like adipose tissue, through an increase in UCP1 and other thermogenic genes [42]. Indeed, mice deficient in FGF21 display diminished browning of WAT [42]. HO-1 upregulation increases PGC-1α, a key player and nuclear regulator of energy metabolism in the heart, liver and kidney, by upregulating mitochondrial biogenesis, promoting β-fatty acid oxidation [28, 43] and upregulating UCP-1 [44]. The upregulation of UCP-1 and other thermogenic genes emphasizes beige adipocytes' key role in thermogenic regulation [4, 45, 46]. Moreover, PGC-1α promotes the differentiation of mesenchymal stem cells into brown-fat adipocytes and prevents the development of metabolic syndrome and T2D and consequent cardiovascular disease [44].
Importantly, we found that adipocyte specific HO-1 expression refurbishes mitochondrial proteins MFN1 and MFN2, in hepatic tissues, all of which are necessary for mitochondrial fusion and functional integrity after ROS-induced damage, thereby triggering the production of beige adipocyte like cells. Increased mitochondrial fusion-to-fission protein ratio allows for increased energy production through its protective role over the ATP gradient, an essential component of respiratory capacity and metabolic activity [47]. It was shown that ablation of adipose HO-1 expression in mice inhibits mitofusion proteins, resulting in increased white fat over beige fat content [39]. In contrast, gene therapy with HO-1 reversed these detrimental effects, and increased mitochondrial fusion over fission. Notably, adipocyte-specific increase in HO-1 reduced adipocyte number and increased mitochondrial biogenesis and function, compared to untreated adipocytes of obese mice. [31, 32, 52, 44, 45].
Another interesting finding is that selective expression of HO-1 in adipocytes positively correlated with decreased hepatic and adipocyte fibrosis in obese mice fed a HFD. We have previously shown that increases in HO-1 in both visceral and subcutaneous fat increase insulin sensitivity [4, 27, 48, 49]. Additionally, the expression of adiponectin is increased by upregulation of HO-1, resulting in improved glucose tolerance and reduced inflammation. In support of these findings, we found increased levels of the insulin receptor (IR-B) and the glucose transporter protein (GLUT-2), evidencing improved insulin sensitivity. Obesity results in decreased adiponectin levels, leading to insulin resistance and hepatic steatosis, and fibrosis [50].
These results agree with several previous findings highlighting the substantial metabolic benefits of pharmacological intervention resulting in HO-1 overexpression [18, 51]. Additionally, we and others demonstrated that the beneficial effect of HO‐1 induction in the prevention of lipid droplet accumulation and fatty liver was reversed by inhibition of HO-1 by HO inhibitors, indicating that HO-1 upregulation was the cause and effect [34, 52]. Thus, low levels of HO‐1 and HO activity are responsible for fatty liver progression. Ablation of adipose-specific HO-1 expression led to an increase in white fat over beige fat through the inhibition of mitochondrial fusion proteins in female mice [39], but deletion of HO-1 in adipocytes had a greater effect on body fat in female mice than in male mice [53]. Our results further support the notion that the improved insulin signaling achieved by targeting adipocyte-specific expression of HO-1 is most likely mediated by increased levels of pAKT and pAMPK.
To test the possibility of potential crosstalk between adipose and hepatic tissue, we previously examined the effects of HO-1 on the adipose tissue derived pro-inflammatory adipokine NOV. Untreated obese mice expressed elevated levels of the Nephroblastoma gene (NOV), increased in obesity and NAFLD, while increased HO-1 expression normalized visceral adipose tissue NOV levels, indicating that induction of HO-1 reprograms white adipose tissue to beige, thereby reducing inflammatory molecules (Figure 4). Increased adiposity and insulin resistance impair liver function and promote the progression of NAFLD to NASH [34, 54, 55]. Moreover, inflammation and impaired antioxidative systems leading to increased ROS production and mitochondrial dysfunction, are implicated in the development of NASH and NAFLD, with the increased risk of liver fibrosis [21].
We have previously shown that pharmacological induction of HO-1 ameliorates NAFLD and NASH in obese mice [34, 38]. In the current study, adipocyte overexpression of HO-1 was sufficient to alleviate liver steatosis and improve the NAS score in obese mice, suggesting a paracrine and/or endocrine influence of HO-1 induction in improving adipocyte function. Additional studies are needed to elucidate the mechanism by which HO-1 expression ameliorates NASH.
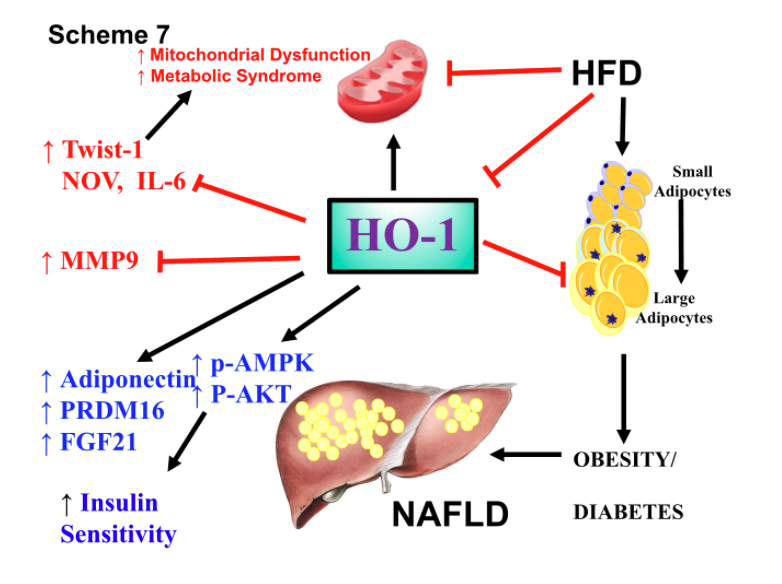
The diagram in Figure 7 summarizes the beneficial effects of elevated levels of adipocyte HO-1 in attenuating NAFLD induced by a high fat diet and restoring mitochondrial function and decreasing inflammation. Some of the possible mechanisms by which adipocyte specific HO-1 expression increases mitochondrial biogenesis and integrity, include inhibition of Fatty acid synthetase (FAS) and subsequent adiposity. HO-1 has been shown to translocate to the nucleus and upregulate transcription factors to reduce insulin resistance and adiposity and increase metabolic function and attenuate NAFLD and obesity. Targeting adipose tissues with specific HO-1 expression attenuates fibrosis, insulin sensitivity and NAFLD in a high-fat diet induced obese murine model.
In conclusion, our data represents a significant contribution to the current understanding of how adipocyte HO-1 functions in ameliorating obesity-associated NAFLD and NASH. NAFLD patients have a much higher incidence of cardiovascular disease and more needs to be done to treat/prevent NAFLD to address this increased risk. Importantly, our findings suggest that increased adipocyte-specific HO-1 expression leads to a reprogramming of white fat stem cells to brown adipocyte-like stem cells, associated with improvement in metabolic function and successfully attenuating the progression of the progression NAFLD to NASH in obesity.
Scoring System for Degree of steatosis (Score 0 ≤ 5%; grade 1 = 0 - 33%; score e 2 = 34%-66%; Score 3 ≥ 66%), lobular inflammation (Score 0: no foci; Score 1: < 2 foci per 200x field; Score 2: 2 to 4 foci per 200x field; Score 3: > 4 foci per 200x field), hepatocyte ballooning (score 0: none; Score 1: rare or few; Score 2: many), and fibrosis (Score 0: no fibrosis; score 1: perisinusoidal or periportal fibrosis; grade 2: perisinusoidal and periportal fibrosis; Score 3: bridging fibrosis; Score 4: cirrhosis).
Table 1. Liver NASH Score
|
NAS pathological score factors |
Control |
HF |
HF + HO-1 |
|
Steatosis |
0 (0.67%) |
2 (49.35%) |
1 (9.28%) |
|
Inflammation |
0 (no foci) |
2 (2-4 foci/field) |
1 (<2 foci/field) |
|
Ballooning |
1 (rare) |
2 (many) |
1 (rare) |
|
Fibrosis |
0 (no fibrosis) |
2 (periportal fibrosis) |
0 (no fibrosis) |
|
NAS value |
1 |
8 NASH |
3 |
Conflicts of Interest
As authors, we declare that we have no conflicts of interest.
Acknowledgements
The authors thank Ms. Jennifer Brown for her editorial assistance.
REFERENCES
1. Hales CM, Carroll MD, Fryar CD, Ogden CL. Prevalence of Obesity and Severe Obesity Among Adults: United States, 2017-2018. NCHS Data Brief. 2020(360):1-8.
2. Andolfi C, Fisichella PM. Epidemiology of Obesity and Associated Comorbidities. Journal of laparoendoscopic & advanced surgical techniques Part A. 2018; 28(8):919-24.
3. Artham SM, Lavie CJ, Milani RV, Ventura HO. Obesity and hypertension, heart failure, and coronary heart disease-risk factor, paradox, and recommendations for weight loss. The Ochsner journal. 2009; 9(3):124-32.
4. Singh SP, Greenberg M, Glick Y, Bellner L, Favero G, Rezzani R, et al. Adipocyte Specific HO-1 Gene Therapy is Effective in Antioxidant Treatment of Insulin Resistance and Vascular Function in an Obese Mice Model. Antioxidants (Basel). 2020; 9(1).
5. Nelson JE, Bhattacharya R, Lindor KD, Chalasani N, Raaka S, Heathcote EJ, et al. HFE C282Y mutations are associated with advanced hepatic fibrosis in Caucasians with nonalcoholic steatohepatitis. Hepatology. 2007; 46(3):723-9.
6. Yan C, Tian X, Li J, Liu D, Ye D, Xie Z, et al. A High-Fat Diet Attenuates AMPK alpha1 in Adipocytes to Induce Exosome Shedding and Nonalcoholic Fatty Liver Development In Vivo. Diabetes. 2021; 70(2):577-88.
7. Vernon G, Baranova A, Younossi ZM. Systematic review: the epidemiology and natural history of non-alcoholic fatty liver disease and non-alcoholic steatohepatitis in adults. Aliment Pharmacol Ther. 2011; 34(3):274-85.
8. Wali JA, Jarzebska N, Raubenheimer D, Simpson SJ, Rodionov RN, O'Sullivan JF. Cardio-Metabolic Effects of High-Fat Diets and Their Underlying Mechanisms-A Narrative Review. Nutrients. 2020; 12(5).
9. Duan Y, Zeng L, Zheng C, Song B, Li F, Kong X, et al. Inflammatory Links Between High Fat Diets and Diseases. Front Immunol. 2018; 9:2649.
10. Speakman JR. Use of high-fat diets to study rodent obesity as a model of human obesity. Int J Obes (Lond). 2019; 43(8):1491-2.
11. Sasson A, Kristoferson E, Batista R, McClung JA, Abraham NG, Peterson SJ. The pivotal role of heme Oxygenase-1 in reversing the pathophysiology and systemic complications of NAFLD. Arch Biochem Biophys. 2021; 697:108679.
12. Cao J, Inoue K, Sodhi K, Puri N, Peterson SJ, Rezzani R, et al. High-fat diet exacerbates renal dysfunction in SHR: reversal by induction of HO-1-adiponectin axis. Obesity (Silver Spring). 2012; 20(5):945-53.
13. Peterson SJ, Vanella L, Gotlinger K, Jiang H, Singh SP, Sodhi K, et al. Oxidized HDL is a potent inducer of adipogenesis and causes activation of the Ang-II and 20-HETE systems in human obese females. Prostaglandins Other Lipid Mediat. 2016; 123:68-77.
14. Lombardi R, Onali S, Thorburn D, Davidson BR, Gurusamy KS, Tsochatzis E. Pharmacological interventions for non-alcohol related fatty liver disease (NAFLD): an attempted network meta-analysis. Cochrane Database Syst Rev. 2017; 3:CD011640.
15. Lafontan M, Girard J. Impact of visceral adipose tissue on liver metabolism Part I: Heterogeneity of adipose tissue and functional properties of visceral adipose tissue. Diabetes Metab. 2008; 34(4):317-27.
16. Donnelly KL, Smith CI, Schwarzenberg SJ, Jessurun J, Boldt MD, Parks EJ. Sources of fatty acids stored in liver and secreted via lipoproteins in patients with nonalcoholic fatty liver disease. J Clin Invest. 2005; 115(5):1343-51.
17. Nicolai A, Li M, Kim DH, Peterson SJ, Vanella L, Positano V, et al. Heme oxygenase-1 induction remodels adipose tissue and improves insulin sensitivity in obesity-induced diabetic rats. Hypertension. 2009; 53(3):508-15.
18. Burgess A, Li M, Vanella L, Kim DH, Rezzani R, Rodella L, et al. Adipocyte heme oxygenase-1 induction attenuates metabolic syndrome in both male and female obese mice. Hypertension. 2010; 56(6):1124-30.
19. Longato L. Non-alcoholic fatty liver disease (NAFLD): a tale of fat and sugar? Fibrogenesis Tissue Repair. 2013; 6(1):14.
20. Chen X, Xu Y, Zhao D, Chen T, Gu C, Yu G, et al. LncRNA-AK012226 Is Involved in Fat Accumulation in db/db Mice Fatty Liver and Non-alcoholic Fatty Liver Disease Cell Model. Front Pharmacol. 2018; 9:888.
21. Chandrasekharan K, Alazawi W. Genetics of Non-Alcoholic Fatty Liver and Cardiovascular Disease: Implications for Therapy? Front Pharmacol. 2019; 10:1413.
22. Fakhouri EW PS, Kothari J, Alex R, Shapiro JI, Abraham NG. Genetic Polymorphisms Complicate COVID-19 Therapy: Pivotal Role of HO-1 in Cytokine Storm. Antioxidants (Basel). 2020.
23. Chang PF, Lin YC, Liu K, Yeh SJ, Ni YH. Heme oxygenase-1 gene promoter polymorphism and the risk of pediatric nonalcoholic fatty liver disease. Int J Obes (Lond). 2015; 39(8):1236-40.
24. Peterson SJ, Rubinstein R, Faroqui M, Raza A, Boumaza I, Zhang Y, et al. Positive Effects of Heme Oxygenase Upregulation on Adiposity and Vascular Dysfunction: Gene Targeting vs. Pharmacologic Therapy. Int J Mol Sci. 2019; 20(10).
25. Sodhi K, Puri N, Favero G, Stevens S, Meadows C, Abraham NG, et al. Fructose Mediated Non-Alcoholic Fatty Liver Is Attenuated by HO-1-SIRT1 Module in Murine Hepatocytes and Mice Fed a High Fructose Diet. PLoS One. 2015; 10(6):e0128648.
26. Sodhi K, Srikanthan K, Goguet-Rubio P, Nichols A, Mallick A, Nawab A, et al. pNaKtide Attenuates Steatohepatitis and Atherosclerosis by Blocking Na/K-ATPase/ROS Amplification in C57Bl6 and ApoE Knockout Mice Fed a Western Diet. Sci Rep. 2017; 7(1):193.
27. Bellner L, Lebovics NB, Rubinstein R, Buchen YD, Sinatra E, Sinatra G, et al. Heme Oxygenase-1 Upregulation: A Novel Approach in the Treatment of Cardiovascular Disease. Antioxidants & redox signaling. 2020; 32(14):1045-60.
28. Singh S, McClung J, Thompson E, Glick Y, Greenberg M, Acosta-Baez G, et al. Cardioprotective heme oxygenase-1-PGC-1α signaling in epicardial fat attenuates cardiovascular risk in humans as in obese mice. Obesity (Silver Spring). 2019.
29. Kawashima A, Oda Y, Yachie A, Koizumi S, Nakanishi I. Heme oxygenase-1 deficiency: the first autopsy case. Hum Pathol. 2002; 33(1):125-30.
30. Radhakrishnan N, Yadav SP, Sachdeva A, Pruthi PK, Sawhney S, Piplani T, et al. Human heme oxygenase-1 deficiency presenting with hemolysis, nephritis, and asplenia. J Pediatr Hematol Oncol. 2011; 33(1):74-8.
31. Poss KD, Tonegawa S. Heme oxygenase 1 is required for mammalian iron reutilization. Proc Natl Acad Sci U S A. 1997; 94:10919-24.
32. Drummond GS, Baum J, Greenberg M, Lewis D, Abraham NG. HO-1 Overexpression and Underexpression: Clinical Implications. Clinical implications Archives of Biochemistry and Biophysics. 2019; 2019 Sep 30; 673:108073.
33. Raffaele M, Carota G, Sferrazzo G, Licari M, Barbagallo I, Sorrenti V, et al. Inhibition of Heme Oxygenase Antioxidant Activity Exacerbates Hepatic Steatosis and Fibrosis In Vitro. Antioxidants (Basel). 2019; 8(8).
34. Sacerdoti D, Singh SP, Schragenheim J, Bellner L, Vanella L, Raffaele M, et al. Development of NASH in Obese Mice is Confounded by Adipose Tissue Increase in Inflammatory NOV and Oxidative Stress. Int J Hepatol. 2018; 2018:3484107.
35. Stacchiotti A, Favero G, Lavazza A, Golic I, Aleksic M, Korac A, et al. Hepatic Macrosteatosis Is Partially Converted to Microsteatosis by Melatonin Supplementation in ob/ob Mice Non-Alcoholic Fatty Liver Disease. PLoS One. 2016; 11(1):e0148115.
36. Matsumoto M, Hada N, Sakamaki Y, Uno A, Shiga T, Tanaka C, et al. An improved mouse model that rapidly develops fibrosis in non-alcoholic steatohepatitis. Int J Exp Pathol. 2013; 94(2):93-103.
37. Schragenheim J, Bellner L, Cao J, Singh SP, Bamshad D, McClung JA, et al. EET enhances renal function in obese mice resulting in restoration of HO-1-Mfn1/2 signaling, and decrease in hypertension through inhibition of sodium chloride co-transporter. Prostaglandins Other Lipid Mediat. 2018; 137:30-9.
38. Raffaele M, Bellner L, Singh SP, Favero G, Rezzani R, Rodella LF, et al. Epoxyeicosatrienoic intervention improves NAFLD in leptin receptor deficient mice by an increase in PGC1alpha-HO-1-PGC1alpha-mitochondrial signaling. Experimental cell research. 2019; 380(2):180-7.
39. Singh S, Grant I, Meissner A, Kappas A, Abraham N. Ablation of adipose-HO-1 expression increases white fat over beige fat through inhibition of mitochondrial fusion and of PGC1alpha in female mice. Horm Mol Biol Clin Investig. 2017; 31(1).
40. Shen HH, Peterson SJ, Bellner L, Choudhary A, Levy L, Gancz L, et al. Cold-Pressed Nigella Sativa Oil Standardized to 3% Thymoquinone Potentiates Omega-3 Protection against Obesity-Induced Oxidative Stress, Inflammation, and Markers of Insulin Resistance Accompanied with Conversion of White to Beige Fat in Mice. Antioxidants (Basel, Switzerland). 2020; 9(6).
41. Jeremic N, Chaturvedi P, Tyagi SC. Browning of White Fat: Novel Insight Into Factors, Mechanisms, and Therapeutics. J Cell Physiol. 2017; 232(1):61-8.
42. Fisher FM, Kleiner S, Douris N, Fox EC, Mepani RJ, Verdeguer F, et al. FGF21 regulates PGC-1alpha and browning of white adipose tissues in adaptive thermogenesis. Genes Dev. 2012; 26(3):271-81.
43. Singh SP, Schragenheim J, Cao J, Falck JR, Abraham NG, Bellner L. PGC-1 alpha regulates HO-1 expression, mitochondrial dynamics and biogenesis: Role of epoxyeicosatrienoic acid. Prostaglandins Other Lipid Mediat. 2016; 125:8-18.
44. Bostrom P, Wu J, Jedrychowski MP, Korde A, Ye L, Lo JC, et al. A PGC1-alpha-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature. 2012; 481(7382):463-8.
45. Cohen P, Spiegelman BM. Brown and Beige Fat: Molecular Parts of a Thermogenic Machine. Diabetes. 2015; 64(7):2346-51.
46. Hattori K, Naguro I, Okabe K, Funatsu T, Furutani S, Takeda K, et al. ASK1 signalling regulates brown and beige adipocyte function. Nat Commun. 2016; 7:11158.
47. Westermann B. Bioenergetic role of mitochondrial fusion and fission. Biochim Biophys Acta. 2012; 1817(10):1833-8.
48. Li M, Kim DH, Tsenovoy PL, Peterson SJ, Rezzani R, Rodella LF, et al. Treatment of obese diabetic mice with a heme oxygenase inducer reduces visceral and subcutaneous adiposity, increases adiponectin levels, and improves insulin sensitivity and glucose tolerance. Diabetes. 2008; 57(6):1526-35.
49. Shen HH, Alex R, Bellner L, Raffaele M, Licari M, Vanella L, et al. Milk thistle seed cold press oil attenuates markers of the metabolic syndrome in a mouse model of dietary-induced obesity. J Food Biochem. 2020:e13522.
50. Polyzos SA, Kountouras J, Zavos C, Tsiaousi E. The role of adiponectin in the pathogenesis and treatment of non-alcoholic fatty liver disease. Diabetes Obes Metab. 2010; 12(5):365-83.
51. Li M, Peterson S, Husney D, Inaba M, Guo K, Kappas A, et al. Long-lasting expression of HO-1 delays progression of type I diabetes in NOD mice. Cell Cycle. 2007; 6(5):567-71.
52. Hinds TD, Jr., Sodhi K, Meadows C, Fedorova L, Puri N, Kim DH, et al. Increased HO-1 levels ameliorate fatty liver development through a reduction of heme and recruitment of FGF21. Obesity (Silver Spring). 2014; 22(3):705-12.
53. Hosick PA, Weeks MF, Hankins MW, Moore KH, Stec DE. Sex-Dependent Effects of HO-1 Deletion from Adipocytes in Mice. Int J Mol Sci. 2017; 18(3).
54. Xia MF, Bian H, Gao X. NAFLD and Diabetes: Two Sides of the Same Coin? Rationale for Gene-Based Personalized NAFLD Treatment. Front Pharmacol. 2019; 10:877.
55. Peng JH, Leng J, Tian HJ, Yang T, Fang Y, Feng Q, et al. Geniposide and Chlorogenic Acid Combination Ameliorates Non-alcoholic Steatohepatitis Involving the Protection on the Gut Barrier Function in Mouse Induced by High-Fat Diet. Front Pharmacol. 2018; 9:1399.
PEER REVIEW
Not commissioned. Externally peer reviewed.
FIGURES
Figure 1. Effects of specific HO-1 expression on adipocytes in lean vs HFD-mice Representative Western blot and densitometry quantification for adipocyte HO-1 levels (A, B) when compared to HFD group. This shows that the mechanism was HO-1 upregulation in adipocytes as well as in hepatocytes.∗p<0.05 versus Lean. #p<0.05 versus HFD. n=5
Figure 2. Adipo-HO-1 upregulation reduces hepatic lipid droplets and fibrosis. (A-C): Morphological analyses of lipid droplet diameter and fibrosis in Lean, HFD-fed and HFD-Adipo-HO-1 groups. Representative Masson’s trichrome stained sections of liver and corresponding morphological analyses are shown. Bar: 40 μm; n= five animals per group. Panel D-E shows representative images of MMP-9 in liver tissues of Lean, HFD and HFD+HO-1 mice group. ∗p<0.05 versus Lean. #p<0.05 versus HFD. n=5
Figure 3. HO-1 upregulation reduces glucose intolerance and hepatic dysfunction. Ln-Adipo-HO-1 overexpression shows remarkable improvements on overall body functions in HFD-fed mice when compared to non-treated HFD fed mice. Data analysis of lenti-adipo-HO-1 treated mice shows improved area under the curve glucose vs. HFD mice (A ) when compared to HFD group. The bar graphs summarize the serum activity of liver-function related enzymes ALT (C), and AST (B). ∗p<0.05 versus Lean. #p<0.05 versus HFD. n=5
Figure 4. HO-1 upregulation increases key genes related to mitochondrial fusion and thermogenesis. mRNA expression levels of mitochondrial thermogenic genes Prdm16 (A), Ucp-1 (B), adiponectin (C) and FGF21(D) in adipose tissue. N= five animals per group. ∗p<0.05 versus Lean. #p<0.05 versus HFD.
Figure 5. Increased HO-1 expression enhances protein expression related to mitochondrial biogenesis and thermogenesis in liver tissue. Representative Western blots of SIRT1, CREG1 and FGF21 and their corresponding densitometry analysis reflecting changes from HO-1 upregulation (A-C); MFN2 and OPA1 (D-F) in liver tissue reflecting changes from HO-1 upregulation. N= five animals per group. ∗p<0.05 versus Lean. #p<0.05 versus HFD.
Figure 6. Adipocyte HO-1 overexpression enhances AMPK, ACC and AKT phosphorylation as well as insulin sensitivity in liver tissue to improve the metabolic syndrome. Panel A and B show representative Western blots and their densitometry analysis for p-AMPK to AMPK, p-ACC to ACC, p-AKT to AKT ratio in the liver tissue. Panel C and D shown representative Western blots and their densitometry analysis for IR-Beta, and GLUT2 in liver tissue, all increased in the treated group. ∗p<0.05 versus Lean. #p<0.05 versus HFD. n=5
Figure 7. Schematic representation of the possible mechanisms by which adipocyte specific HO-1 expression increases mitochondrial biogenesis and integrity, which inhibits Fatty acid synthetase (FAS) and adiposity. HO-1 has been shown to translocate to the nucleus and upregulate transcription factors to reduce insulin resistance and adiposity and increase metabolic function and attenuates NAFLD and obesity. Targeting adipose tissues with specific HO-1 expression attenuates fibrosis, insulin sensitivity and NAFLD in an obese mice model.
1. Hales CM, Carroll MD, Fryar CD, Ogden CL. Prevalence of Obesity and Severe Obesity Among Adults: United States, 2017-2018. NCHS Data Brief. 2020(360):1-8.
2. Andolfi C, Fisichella PM. Epidemiology of Obesity and Associated Comorbidities. Journal of laparoendoscopic & advanced surgical techniques Part A. 2018; 28(8):919-24.
3. Artham SM, Lavie CJ, Milani RV, Ventura HO. Obesity and hypertension, heart failure, and coronary heart disease-risk factor, paradox, and recommendations for weight loss. The Ochsner journal. 2009; 9(3):124-32.
4. Singh SP, Greenberg M, Glick Y, Bellner L, Favero G, Rezzani R, et al. Adipocyte Specific HO-1 Gene Therapy is Effective in Antioxidant Treatment of Insulin Resistance and Vascular Function in an Obese Mice Model. Antioxidants (Basel). 2020; 9(1).
5. Nelson JE, Bhattacharya R, Lindor KD, Chalasani N, Raaka S, Heathcote EJ, et al. HFE C282Y mutations are associated with advanced hepatic fibrosis in Caucasians with nonalcoholic steatohepatitis. Hepatology. 2007; 46(3):723-9.
6. Yan C, Tian X, Li J, Liu D, Ye D, Xie Z, et al. A High-Fat Diet Attenuates AMPK alpha1 in Adipocytes to Induce Exosome Shedding and Nonalcoholic Fatty Liver Development In Vivo. Diabetes. 2021; 70(2):577-88.
7. Vernon G, Baranova A, Younossi ZM. Systematic review: the epidemiology and natural history of non-alcoholic fatty liver disease and non-alcoholic steatohepatitis in adults. Aliment Pharmacol Ther. 2011; 34(3):274-85.
8. Wali JA, Jarzebska N, Raubenheimer D, Simpson SJ, Rodionov RN, O'Sullivan JF. Cardio-Metabolic Effects of High-Fat Diets and Their Underlying Mechanisms-A Narrative Review. Nutrients. 2020; 12(5).
9. Duan Y, Zeng L, Zheng C, Song B, Li F, Kong X, et al. Inflammatory Links Between High Fat Diets and Diseases. Front Immunol. 2018; 9:2649.
10. Speakman JR. Use of high-fat diets to study rodent obesity as a model of human obesity. Int J Obes (Lond). 2019; 43(8):1491-2.
11. Sasson A, Kristoferson E, Batista R, McClung JA, Abraham NG, Peterson SJ. The pivotal role of heme Oxygenase-1 in reversing the pathophysiology and systemic complications of NAFLD. Arch Biochem Biophys. 2021; 697:108679.
12. Cao J, Inoue K, Sodhi K, Puri N, Peterson SJ, Rezzani R, et al. High-fat diet exacerbates renal dysfunction in SHR: reversal by induction of HO-1-adiponectin axis. Obesity (Silver Spring). 2012; 20(5):945-53.
13. Peterson SJ, Vanella L, Gotlinger K, Jiang H, Singh SP, Sodhi K, et al. Oxidized HDL is a potent inducer of adipogenesis and causes activation of the Ang-II and 20-HETE systems in human obese females. Prostaglandins Other Lipid Mediat. 2016; 123:68-77.
14. Lombardi R, Onali S, Thorburn D, Davidson BR, Gurusamy KS, Tsochatzis E. Pharmacological interventions for non-alcohol related fatty liver disease (NAFLD): an attempted network meta-analysis. Cochrane Database Syst Rev. 2017; 3:CD011640.
15. Lafontan M, Girard J. Impact of visceral adipose tissue on liver metabolism Part I: Heterogeneity of adipose tissue and functional properties of visceral adipose tissue. Diabetes Metab. 2008; 34(4):317-27.
16. Donnelly KL, Smith CI, Schwarzenberg SJ, Jessurun J, Boldt MD, Parks EJ. Sources of fatty acids stored in liver and secreted via lipoproteins in patients with nonalcoholic fatty liver disease. J Clin Invest. 2005; 115(5):1343-51.
17. Nicolai A, Li M, Kim DH, Peterson SJ, Vanella L, Positano V, et al. Heme oxygenase-1 induction remodels adipose tissue and improves insulin sensitivity in obesity-induced diabetic rats. Hypertension. 2009; 53(3):508-15.
18. Burgess A, Li M, Vanella L, Kim DH, Rezzani R, Rodella L, et al. Adipocyte heme oxygenase-1 induction attenuates metabolic syndrome in both male and female obese mice. Hypertension. 2010; 56(6):1124-30.
19. Longato L. Non-alcoholic fatty liver disease (NAFLD): a tale of fat and sugar? Fibrogenesis Tissue Repair. 2013; 6(1):14.
20. Chen X, Xu Y, Zhao D, Chen T, Gu C, Yu G, et al. LncRNA-AK012226 Is Involved in Fat Accumulation in db/db Mice Fatty Liver and Non-alcoholic Fatty Liver Disease Cell Model. Front Pharmacol. 2018; 9:888.
21. Chandrasekharan K, Alazawi W. Genetics of Non-Alcoholic Fatty Liver and Cardiovascular Disease: Implications for Therapy? Front Pharmacol. 2019; 10:1413.
22. Fakhouri EW PS, Kothari J, Alex R, Shapiro JI, Abraham NG. Genetic Polymorphisms Complicate COVID-19 Therapy: Pivotal Role of HO-1 in Cytokine Storm. Antioxidants (Basel). 2020.
23. Chang PF, Lin YC, Liu K, Yeh SJ, Ni YH. Heme oxygenase-1 gene promoter polymorphism and the risk of pediatric nonalcoholic fatty liver disease. Int J Obes (Lond). 2015; 39(8):1236-40.
24. Peterson SJ, Rubinstein R, Faroqui M, Raza A, Boumaza I, Zhang Y, et al. Positive Effects of Heme Oxygenase Upregulation on Adiposity and Vascular Dysfunction: Gene Targeting vs. Pharmacologic Therapy. Int J Mol Sci. 2019; 20(10).
25. Sodhi K, Puri N, Favero G, Stevens S, Meadows C, Abraham NG, et al. Fructose Mediated Non-Alcoholic Fatty Liver Is Attenuated by HO-1-SIRT1 Module in Murine Hepatocytes and Mice Fed a High Fructose Diet. PLoS One. 2015; 10(6):e0128648.
26. Sodhi K, Srikanthan K, Goguet-Rubio P, Nichols A, Mallick A, Nawab A, et al. pNaKtide Attenuates Steatohepatitis and Atherosclerosis by Blocking Na/K-ATPase/ROS Amplification in C57Bl6 and ApoE Knockout Mice Fed a Western Diet. Sci Rep. 2017; 7(1):193.
27. Bellner L, Lebovics NB, Rubinstein R, Buchen YD, Sinatra E, Sinatra G, et al. Heme Oxygenase-1 Upregulation: A Novel Approach in the Treatment of Cardiovascular Disease. Antioxidants & redox signaling. 2020; 32(14):1045-60.
28. Singh S, McClung J, Thompson E, Glick Y, Greenberg M, Acosta-Baez G, et al. Cardioprotective heme oxygenase-1-PGC-1α signaling in epicardial fat attenuates cardiovascular risk in humans as in obese mice. Obesity (Silver Spring). 2019.
29. Kawashima A, Oda Y, Yachie A, Koizumi S, Nakanishi I. Heme oxygenase-1 deficiency: the first autopsy case. Hum Pathol. 2002; 33(1):125-30.
30. Radhakrishnan N, Yadav SP, Sachdeva A, Pruthi PK, Sawhney S, Piplani T, et al. Human heme oxygenase-1 deficiency presenting with hemolysis, nephritis, and asplenia. J Pediatr Hematol Oncol. 2011; 33(1):74-8.
31. Poss KD, Tonegawa S. Heme oxygenase 1 is required for mammalian iron reutilization. Proc Natl Acad Sci U S A. 1997; 94:10919-24.
32. Drummond GS, Baum J, Greenberg M, Lewis D, Abraham NG. HO-1 Overexpression and Underexpression: Clinical Implications. Clinical implications Archives of Biochemistry and Biophysics. 2019; 2019 Sep 30; 673:108073.
33. Raffaele M, Carota G, Sferrazzo G, Licari M, Barbagallo I, Sorrenti V, et al. Inhibition of Heme Oxygenase Antioxidant Activity Exacerbates Hepatic Steatosis and Fibrosis In Vitro. Antioxidants (Basel). 2019; 8(8).
34. Sacerdoti D, Singh SP, Schragenheim J, Bellner L, Vanella L, Raffaele M, et al. Development of NASH in Obese Mice is Confounded by Adipose Tissue Increase in Inflammatory NOV and Oxidative Stress. Int J Hepatol. 2018; 2018:3484107.
35. Stacchiotti A, Favero G, Lavazza A, Golic I, Aleksic M, Korac A, et al. Hepatic Macrosteatosis Is Partially Converted to Microsteatosis by Melatonin Supplementation in ob/ob Mice Non-Alcoholic Fatty Liver Disease. PLoS One. 2016; 11(1):e0148115.
36. Matsumoto M, Hada N, Sakamaki Y, Uno A, Shiga T, Tanaka C, et al. An improved mouse model that rapidly develops fibrosis in non-alcoholic steatohepatitis. Int J Exp Pathol. 2013; 94(2):93-103.
37. Schragenheim J, Bellner L, Cao J, Singh SP, Bamshad D, McClung JA, et al. EET enhances renal function in obese mice resulting in restoration of HO-1-Mfn1/2 signaling, and decrease in hypertension through inhibition of sodium chloride co-transporter. Prostaglandins Other Lipid Mediat. 2018; 137:30-9.
38. Raffaele M, Bellner L, Singh SP, Favero G, Rezzani R, Rodella LF, et al. Epoxyeicosatrienoic intervention improves NAFLD in leptin receptor deficient mice by an increase in PGC1alpha-HO-1-PGC1alpha-mitochondrial signaling. Experimental cell research. 2019; 380(2):180-7.
39. Singh S, Grant I, Meissner A, Kappas A, Abraham N. Ablation of adipose-HO-1 expression increases white fat over beige fat through inhibition of mitochondrial fusion and of PGC1alpha in female mice. Horm Mol Biol Clin Investig. 2017; 31(1).
40. Shen HH, Peterson SJ, Bellner L, Choudhary A, Levy L, Gancz L, et al. Cold-Pressed Nigella Sativa Oil Standardized to 3% Thymoquinone Potentiates Omega-3 Protection against Obesity-Induced Oxidative Stress, Inflammation, and Markers of Insulin Resistance Accompanied with Conversion of White to Beige Fat in Mice. Antioxidants (Basel, Switzerland). 2020; 9(6).
41. Jeremic N, Chaturvedi P, Tyagi SC. Browning of White Fat: Novel Insight Into Factors, Mechanisms, and Therapeutics. J Cell Physiol. 2017; 232(1):61-8.
42. Fisher FM, Kleiner S, Douris N, Fox EC, Mepani RJ, Verdeguer F, et al. FGF21 regulates PGC-1alpha and browning of white adipose tissues in adaptive thermogenesis. Genes Dev. 2012; 26(3):271-81.
43. Singh SP, Schragenheim J, Cao J, Falck JR, Abraham NG, Bellner L. PGC-1 alpha regulates HO-1 expression, mitochondrial dynamics and biogenesis: Role of epoxyeicosatrienoic acid. Prostaglandins Other Lipid Mediat. 2016; 125:8-18.
44. Bostrom P, Wu J, Jedrychowski MP, Korde A, Ye L, Lo JC, et al. A PGC1-alpha-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature. 2012; 481(7382):463-8.
45. Cohen P, Spiegelman BM. Brown and Beige Fat: Molecular Parts of a Thermogenic Machine. Diabetes. 2015; 64(7):2346-51.
46. Hattori K, Naguro I, Okabe K, Funatsu T, Furutani S, Takeda K, et al. ASK1 signalling regulates brown and beige adipocyte function. Nat Commun. 2016; 7:11158.
47. Westermann B. Bioenergetic role of mitochondrial fusion and fission. Biochim Biophys Acta. 2012; 1817(10):1833-8.
48. Li M, Kim DH, Tsenovoy PL, Peterson SJ, Rezzani R, Rodella LF, et al. Treatment of obese diabetic mice with a heme oxygenase inducer reduces visceral and subcutaneous adiposity, increases adiponectin levels, and improves insulin sensitivity and glucose tolerance. Diabetes. 2008; 57(6):1526-35.
49. Shen HH, Alex R, Bellner L, Raffaele M, Licari M, Vanella L, et al. Milk thistle seed cold press oil attenuates markers of the metabolic syndrome in a mouse model of dietary-induced obesity. J Food Biochem. 2020:e13522.
50. Polyzos SA, Kountouras J, Zavos C, Tsiaousi E. The role of adiponectin in the pathogenesis and treatment of non-alcoholic fatty liver disease. Diabetes Obes Metab. 2010; 12(5):365-83.
51. Li M, Peterson S, Husney D, Inaba M, Guo K, Kappas A, et al. Long-lasting expression of HO-1 delays progression of type I diabetes in NOD mice. Cell Cycle. 2007; 6(5):567-71.
52. Hinds TD, Jr., Sodhi K, Meadows C, Fedorova L, Puri N, Kim DH, et al. Increased HO-1 levels ameliorate fatty liver development through a reduction of heme and recruitment of FGF21. Obesity (Silver Spring). 2014; 22(3):705-12.
53. Hosick PA, Weeks MF, Hankins MW, Moore KH, Stec DE. Sex-Dependent Effects of HO-1 Deletion from Adipocytes in Mice. Int J Mol Sci. 2017; 18(3).
54. Xia MF, Bian H, Gao X. NAFLD and Diabetes: Two Sides of the Same Coin? Rationale for Gene-Based Personalized NAFLD Treatment. Front Pharmacol. 2019; 10:877.
55. Peng JH, Leng J, Tian HJ, Yang T, Fang Y, Feng Q, et al. Geniposide and Chlorogenic Acid Combination Ameliorates Non-alcoholic Steatohepatitis Involving the Protection on the Gut Barrier Function in Mouse Induced by High-Fat Diet. Front Pharmacol. 2018; 9:1399.
The Roles of Choline in Maintaining Optimal Health
The Effects of Intake of Bread with Treated Corn Bran Inclusion on Postprandial Glyceamic Response
Food Waste throughout the Food Production Continuum – Water Food and Energy Nexus
Use of Lightly Potassium-Enriched Soy Sauce at Home Reduced Urinary Sodium-to-Potassium Ratio
A Different Type of Critical Migration
PRECEDE: A Conceptual Model to Assess Immigrant Health
Walking Together: Supporting Indigenous Student Success in University
Cosmetic Surgery and Body Image in Race/Ethnic Minorities

Our articles most useful
Adrian Taylor and Marica Bakovic*
Published : June 21, 2019
Journal of Food & Nutritional Sciences
Zhimin Cui, Lynne Kennedy, Weili Li*
Published : September 30, 2019
Journal of Food & Nutritional Sciences
Elena Castell-Perez*, Rosana G. Moreira, Hal S. Knowles, III
Published : October 01, 2019
Journal of Food & Nutritional Sciences
Nagako Okuda, Makoto Miura, Kazuyoshi Itai, Takuya Morikawa, Junko Sasaki, Tamami Asanuma, Mikako Fujii, Akira Okayama
Published : March 27, 2019
Journal of Food & Nutritional Sciences