This study’s aim was to compare the effects of various processing, including non-fermentation and fermentation technologies on polyphenols and antioxidant properties in traditional soy foods by using the same Prosoy soybean cultivar. Soybean was processed into non-fermented products (soymilk by two technologies, and tofu (by two technologies), and fermented products (natto aged for 2, 4, and 6 days; tempeh aged for 2, 4 and 6 days; and soy-yogurt aged for 2, 4, 6, 8 days). Total phenolic content (TPC), total flavonoids content (TFC), condensed tannins content (CTC), individual phenolic acids, isoflavones, DPPH, ferric-reducing antioxidant power (FRAP), and oxygen radical absorbance capacity (ORAC) were measured. Results showed natto had the highest TPC among all products. Cooked soymilk(s) had the highest CTC (1.1-0.9 mg CAE/g) among all products. In general, fermented products showed higher (p <0.05) DPPH and FRAP antioxidant activities compared to non-fermented soy foods. ORAC values of natto and cooked soymilk ranked higher (p<0.05) than all products. Aging did not affect the total phenolic acid content in natto, tempeh or yogurt. Total isoflavone content of soymilk, yogurt, filled tofu (2027-2248 µg/g) was higher (p < 0.05) than natto and tempeh (1552.2-2013.7 µg/g). Significant changes occurred in the phenolic and antioxidant properties of soybean products and the extent of changes was dependent upon processing methods. Fermented products and non-fermented soy foods have unique compositions of isoflavones, phenolic acid compounds and antioxidant activities. The study contributes to new knowledge for soy consumers to choose different types of products for enhancing health, and for the processing industry to optimize products for the consumers.
*Corresponding Author:
Sam K. C. Chang, PhD
Coastal Research and Extension, 3411 Frederic Street
Pascagoula, MS 39567, USA; Tel: +228-762-7783; Fax: +228-388-7493; E-mail: sc1690@msstate.edu
ABSTRACT
This study’s aim was to compare the effects of various processing, including non-fermentation and fermentation technologies on polyphenols and antioxidant properties in traditional soy foods by using the same Prosoy soybean cultivar. Soybean was processed into non-fermented products (soymilk by two technologies, and tofu (by two technologies), and fermented products (natto aged for 2, 4, and 6 days; tempeh aged for 2, 4 and 6 days; and soy-yogurt aged for 2, 4, 6, 8 days). Total phenolic content (TPC), total flavonoids content (TFC), condensed tannins content (CTC), individual phenolic acids, isoflavones, DPPH, ferric-reducing antioxidant power (FRAP), and oxygen radical absorbance capacity (ORAC) were measured. Results showed natto had the highest TPC among all products. Cooked soymilk(s) had the highest CTC (1.1-0.9 mg CAE/g) among all products. In general, fermented products showed higher (p <0.05) DPPH and FRAP antioxidant activities compared to non-fermented soy foods. ORAC values of natto and cooked soymilk ranked higher (p<0.05) than all products. Aging did not affect the total phenolic acid content in natto, tempeh or yogurt. Total isoflavone content of soymilk, yogurt, filled tofu (2027-2248 µg/g) was higher (p < 0.05) than natto and tempeh (1552.2-2013.7 µg/g). Significant changes occurred in the phenolic and antioxidant properties of soybean products and the extent of changes was dependent upon processing methods. Fermented products and non-fermented soy foods have unique compositions of isoflavones, phenolic acid compounds and antioxidant activities. The study contributes to new knowledge for soy consumers to choose different types of products for enhancing health, and for the processing industry to optimize products for the consumers.
Keywords: Phenolic acids, isoflavones, antioxidants, fermentation, natto, tempeh, soybean, soybean sprouts, tofu.
|
|
ABBREVIATIONS
|
Total phenolic content |
TPC |
|
Total flavonoids content |
TFC |
|
Condensed tannins content |
CTC |
|
2,2-Diphenyl-1-picrylhydrazyl |
DPPH |
|
Ferric-reducing antioxidant power |
FRAP |
|
Oxygen radical absorbance capacity |
ORAC |
|
Gallic acid equivalent |
GAE |
|
Trifluoroacetic acid |
TFA |
|
High performance liquid chromatography |
HPLC |
|
2,4,6-tri(2-pyridyl)-s-triazine |
TPTZ |
|
ethylenediaminetetraacetic acid |
EDTA |
|
Association of Official Analytical Chemists |
AOAC |
|
cooked traditional soymilk |
CTSmilk |
|
soymilk cooked with okara |
CSmilkOF |
|
Principal Component Analysis |
PCA |
|
Gallic acid |
GA |
|
Protocatechuic acid |
PA |
|
2,3,4-trihydroxybenzoic acid |
TBA |
|
Protocatechualdehyde |
PCD |
|
p-hydroxybenzoic acid |
HBA |
|
Gentistic acid |
GNA |
|
Syringic acid |
SA |
|
Vanillic acid |
VA |
|
Vanillin |
VN |
|
Chlorogenic acid |
CLA |
|
p-coumaric acid + syringaldehyde |
PCA+SD |
|
m-coumaric acid + ferullic acid |
MCA+FA |
|
Sinapic acid |
SPA |
|
o-coumaric acid |
OCA |
|
raw soymilk slurry |
RSmilk slurry |
|
cooked soymilk with okra |
CSmilkOF |
|
Diadzin |
Din |
|
Genistin |
Gin |
|
Glycitin |
Gly |
|
Malonyldaidzin |
MDin |
|
Malonylgenistin |
MGin |
|
Malonylglycitin |
MGly |
|
Acetyldiadzin |
ADin |
|
Acetylgenistin |
AGin |
|
Acetylglycitin |
AGly |
|
Diadzein |
Dein |
|
Genistein |
Gein |
|
Glycitein |
Glein |
|
subtotal diadzein |
T-Dein |
|
subtotal genistein |
T-Gein |
|
subtotal glycitein |
T-Glein |
|
Total Isoflavones |
TI |
|
Yogurt day 0 |
Yogurt D0 |
|
Tempeh day 2 |
TempD2 |
|
Natto day 2 |
NattoD2 |
|
No detectable |
ND |
INTRODUCTION
Soybean (Glycine max) and its food products, containing phytoestrogen isoflavones and antioxidant polyphenols, have been reported to protect against a variety of diseases [1-4]. Reactive oxygen species cause oxidative stress in most pathological cases and antioxidants play an important role to defend against cellular damage [5]. The combined isoflavone and phenolic acids are responsible for nearly all antioxidant activities in soybean and soy products [2, 6]. The distribution of phenolic compounds in commercial soybean and soy products depends on genetic factors (seed shape, size, color, etc.), environmental factors (temperature, precipitation, storage period and conditions), processing conditions (heating, steaming, grinding, soaking, and fermentation) and dilution with non-soy-ingredient [7-11].
Raw soybeans contain trypsin inhibitor and hemagglutinins and have an unpalatable hard texture and strong objectionable beany odor and must be thermally processed for human consumption. Traditional soy foods are manufactured with different combinations of unit operations of soaking, grinding, filtration, heating, coagulation, pressing, and microbial fermentation or germination. The variations in technologies affect the structures and compositions of phenolic contents and antioxidant activities [7, 8, 10, 12-14]. Our recent research [10] showed that a combination of grinding temperature and heating technologies greatly influenced the isoflavone composition in soymilk, resulting in lower malonylglucosides and higher glucosides and aglycones. Our lab research on black soybean showed that grinding with different ratios of water in two steps, or cycling okara-rewashed water during soymilk production influenced the isoflavone compositions and cancer cell inhibitory capability [8].
Chung and others [15] attributed the low recovery (47.1%) of 43 phenolic compounds in tofu to the boiling, leaching in water, filtering, coagulation, and whey exclusion. Coagulation caused significant loss in total isoflavones during tofu making [7]. Fermentation can release phenolics from soybean as a result of the action of the cell wall-degrading microbial enzymes and through bioconversion of flavonoids into new metabolites [16-18]. Solid-state fermentation yields higher secondary metabolites than submerged fermentation [19]. Rhizopus oligosporus is employed in solid-state fermentation of tempeh. Higher antioxidant activity is linked to α- and β-glucosidase and β-glucuronidase activities, while high total phenolic content linked to α-amylase during fermentation of whole soybean using Rhizopus oligosporus [18]. Soaking and cooking caused significant losses of phenolic content, while fermentation replenished the total phenol to the level present in raw soybean during tempeh manufacture [20]. Lactic acid bacteria with β-glucosidase activity increased the isoflavone aglycone content in fermented soymilk [21, 22]. Bacillus subtilis can enhance aglycones, malonylglucosides, gallic acid, flavanols, vitamin K2, succinyl genistin and succinyl diadzin [23] in Japanese natto (3), or Korean soybean paste cheonggukjang [24] or Thailand thua nao [25]. Toda and others [26] investigated the isoflavone composition of 50 commercial soy foods consumed in Japan and reported that the heating method and temperature was the primary factors in clustering soy-products. Tofu, produced with mild heating (85-105°C for a short time) conditions showed similar isoflavone distribution as raw soybean, while soymilk and natto produced with comparatively stronger heat (pressurized steaming) contained lower proportion of malonylglucosides and higher β-glucosides.
Studies from our laboratory and others have reported the individual phenolic compounds, total phenol or antioxidant activities in soybean seed or processed/fermented soy foods [9, 15, 18, 20, 26-28]. Due to the wide differences in soybean materials and processing methodologies used by different laboratories, it is difficult to make a fair comparison of processing effect on the extent of phenolic and antioxidant changes in various soy products. To the best of our knowledge, a comprehensive and systematic comparison of different processing techniques across major soy food products manufactured from the same soybean variety is lacking. Therefore, our objective was to investigate the relative changes of phenolic compounds, isoflavones, and antioxidant properties as affected by processing methods commonly used for making traditional soy foods made from one single soybean cultivar.
MATERIALS AND METHODS
Materials
The high-protein Prosoy cultivar, a yellow color food-grade soybean, was obtained from SB&B Co. (Fargo, ND, U.S.A.) and stored at -20°C until use.
Chemicals
Sixteen phenolic acids, three aldehydes, HPLC-grade trifluoroacetic acid (TFA), 2-diphenyl-1-picrylhydrazyl radical (DPPH), Folin-Ciocalteu reagent, 2,4,6-tri(2-pyridyl)-s-triazine (TPTZ), and 6-hydroxy-2,5,7,8-tetramethyl chroman-2-carboxylic acid (Trolox), 2,2′-Azobis(2-amidinopropane) dihydrochloride (AAPH), isoflavones (daidzin, genistin, glycitin, daidzein, genistein, glycitein, apigenin), and other chemicals, including HPLC and analytical grade solvents has been reported in our study [12].
Production of soybean products
Soymilk
Two kinds of soymilk were prepared, the first kind was traditional soymilk (Chinese processing style) prepared according to our laboratory method [29] while the second method (Japanese style) used soy slurry for heating followed by filtration. In brief, for the Chinese style method, in which the insoluble residues (okara) was filtered out to obtain residue-free raw soymilk prior to cooking, 300 g of soybeans were soaked in 1500 mL ambient temperature water for 16 h before grinding (Oster blender model 4918, Boca Raton, FL) with bean-to-water ratio of 1:9 (w/w). The soy slurry was filtered to remove okara using muslin cloth, and resulted soymilk heated was boiled for 20 min to remove trypsin inhibitors [8, 9]. The second kind of soymilk was processed by a Japanese style: The soy slurry after grinding was boiled for 20 min, followed by filtration through muslin cloth to separate the okara from the soymilk. Each type of soymilk was prepared in triplicate and cooled in an ice bath before freeze-drying.
Tofu
Pressed tofu was prepared as described in our recent publication [30]. Briefly, for pressed tofu making, soybean was soaked, ground in water and the slurry was filtered. The obtained raw soymilk was boiled for 5 min and cooled to 85°C, at which CaSO4 at 2% raw bean weight was added to coagulate the hot soymilk to form curd, which was broken and pressed in a wooden box to release whey. After pressing, tofu was freeze-dried for further analyses.
Filled tofu were prepared also as described in our recent paper [30]. Briefly, soybean was soaked, ground in water and the slurry was filtered to obtain raw soymilk. The soymilk was cooked to 90°C to denature proteins; and the heated soymilk was cooled to 4°C and mixed with MgCl2 as a coagulant with a final concentration of 0.25%. The cold soymilk containing coagulant in a container was heated at 85°C for 45 min to form tofu without breaking the curd so the whey was retained in the tofu. The filled tofu was freeze-dried for analysis.
Natto
Natto was prepared following our laboratory method. Bacillus natto culture was prepared from a commercial natto product purchased from Asian grocery store in Mississippi. About 10 g of commercial natto were kept in 200 mL 8% Difco Nutrient broth (BD Diagnostics, Sparks, MD) and incubated at 40°C for 16 h on a shaker to get OD660 = 1.5. The bacterial broth was centrifuged at 15000 g for 25 min to remove residues and diluted with 20 mL sterile water. About 50 g of soybeans were soaked at room temperature for 16 h, followed by sterilizing in aluminum foil at 121°C for 35 min. The sterilized bean was placed in a styrofoam sandwich box with holes in the cover and inoculated with 5 mL bacterial suspension. The beans were incubated in an incubator at 40°C for 19 h. All bacterial transfer experiments were conducted under aseptic conditions under a laminar flow hood. Samples were prepared in triplicate before freeze-drying.
Tempeh
Tempeh was prepared using starter culture, Rhizopus oryzae obtained from the Cultures for Health (Morrisville, NC). The method for making tempeh was adopted from the same company with minor modifications. In brief, about 300 g of soybeans were boiled with tap water with bean-to-water ratio 1:4 (w/w) for 1 h until cotyledons became hydrated. Beans were dehulled and cleaned, followed by soaking in the previously boiled water at 30°C till pH reached 6.5. The soaked beans were washed and steamed for 15 min. The beans were dried on low heat in a pot to evaporate the water off the beans. Tempeh starter was added into the dried soybean with soybean to tempeh starter ratio of 100 g to 0.2 g. The inoculated beans were mixed and packed in plastic zip lock bags with pores and incubated at 31°C for 48 h on cooling racks to facilitate fungi to grow all over the surface as evidence of white mycelia covering the entire beans. Tempeh were kept at 4°C for 2, 4, and 6 days before freeze-drying.
Soy yogurt
Soymilk was made using the hot grinding method developed in our laboratory [29]. In brief, 300g of soybeans were soaked in 1500 mL tap water for 16 h followed by grinding with tap water with a bean-to-water ratio of 1:7 (w/w). Soymilk was heated according to the traditional stove cooking method [9]. Set-style soy yogurt was prepared according Donkor and others [31]. Soymilk was sterilized at 121°C for 5 min in glass bottles and allowed to cool to 40°C [32]. The cooled soymilk was aseptically inoculated, at the manufacturer’s recommended rate, with a yogurt culture (YC-087; Chr. Hansen Laboratory, Inc., Milwaukee, WI) that contained strains of Streptococcus thermophilus and Lactobacillus delbrueckii ssp. Bulgaricus. The inoculated soymilk batches were poured into 500 mL sterile clear/transparent plastic cups with lids (300 mL per cup) and incubated at 42°C until the pH of 4.5 was reached. The products were removed from the incubator and stored at 4°C for up to 8 days before sampling on days 0, 2, 4, 6, and 8 and freeze-drying.
Grinding and extraction of total phenolics from soybean products
Grinding and phenolic extraction were conducted according to the method described in our previous study [33]. Freeze-dried products were ground with a coffee grinder (KitchenAid, Benton Harbor, MI) and passed through a 60-mesh sieve. The powder was stored in an air-tight container at -20°C until use. The extract was prepared using 0.5 g of powdered sample and 10 mL acetone/water (50:50, v/v) as described in our previous study [12]. The extract was stored at -20°C in the dark until further assays.
Determination of phenolic substances and antioxidant properties
Determination of total phenolic content (TPC), total flavonoid content (TFC), condensed tannin content (CTC), DPPH scavenging activity (DPPH), ferric reducing antioxidant power (FRAP), oxygen radical absorbing capacity (ORAC)
The TPC, TFC, CTC, DPPH, FRAP were determined using the colorimetric method modified in our laboratory [14]. TPC was expressed as milligrams of gallic acid equivalents (mg of GAE/g of sample) through the calibration curve of gallic acid. The TFC was expressed as milligrams of catechin equivalents (mg of CAE/g of sample) using the calibration curve of (+)-catechin. The amount of condensed tannin was expressed as milligrams of catechin equivalents (mg of CAE/g of sample) using the calibration curve of
(+)- catechin. DPPH was expressed as micromoles of Trolox equivalents per gram of sample (mmol TE/g). FRAP values were expressed as millimoles of Fe2+ equivalents per 100g of sample (mmol Fe2+ equivalents/100 g). ORAC was performed as described by [12] and expressed as micromoles of Trolox equivalents per gram sample (mmol TE/g).
HPLC analysis of phenolic acid content
Extraction of free and total phenolic acids content: The extraction of free and conjugated phenolic acids was performed according to Kumari and Chang [12] after minor modification from Xu and Chang [14]. Free phenolic acids were extracted using methanol/water/acetic acid/butylated hydroxytoluene extraction solvent (85:15:0.5:0.2, v/v/v/v/v), while total phenolic acids were extracted with ethyl acetate after alkaline hydrolysis in the presence of ethylenediaminetetraacetic acid (EDTA) and ascorbic acid. Phenolic acids were expressed as mg/g on a dry weight basis.
HPLC analysis of isoflavone compositions
Extraction and HPLC analysis of isoflavones: Isoflavones were extracted and analyzed using AOAC method 2008.03 (34) as explained in our previous study [12]. The same Agilent technology chromatography system as used for phenolic acid analysis was used for the analysis of isoflavones.
Identification and quantification of isoflavones: The internal calibration curves were obtained for each of the six external standards by plotting calibration standard concentration of each standard against the area ratio of the standard peak to the internal standard peak. The concentrations of malonyl and acetyl forms were calculated from the standard curves of aglycone forms, and then multiplied with conversion factors as described by Collison [34]. The isoflavone contents were expressed as µg of isoflavone per gram on dry weight basis.
Statistical analysis
Each processing was done in triplicate on different dates. Data were subjected to analysis of variance (ANOVA) with SAS 9.4 package (SAS Institute Inc., Cary, NC). Significant differences among variables were determined by Duncan’s multiple range test (p <0.05). Data were expressed as means ± SD (n=3). The multivariate analysis was performed using XLSTAT 20015.04.
RESULTS AND DISCUSSION
Effect of processing on total phenolic content and antioxidant activities
The TPC, TFC, CTC, DPPH, FRAP and ORAC of the extracts from the same original raw Prosoy soybean and processed products, including traditional raw soymilk, traditional cooked soymilk (Chinese style soymilk), raw soymilk slurry, soymilk cooked with okara and filtered (Japanese style soymilk), tofu (filled and pressed), tempeh (2, 4 and 6 day stored at 4°C), natto (2, 4 and 6 day stored at 4°C) and yogurt (0, 2, 4, 6, 8 day stored at 4°C) are presented in Figure 1. In comparison to the raw soybean (about 3 mg/g), TPC increased by 125-48% in natto, by 46-12% in yogurt, and by 56% and 32%, in cooked traditional soymilk (CTSmilk) and soymilk cooked with okara (CSmilkOF), respectively, indicating that the processing technologies for making these products could release or increase phenolic compounds [10, 15, 17]. No significant differences in TPC were observed among filled tofu and tempeh (days 2, 4, 6), ranging between 2.0-3.2 mg GAE/g. Haron and Raob [20] observed 59% and 35% reduction in TPC during soaking and boiling of tempeh, while 65% increases occurred during fermentation to a level not different (p > 0.05) from the original soybean. This was similar to our results since only minor or no differences existed between tempeh and raw soybean. Higher (p <0.05) TFC values were observed in traditionally cooked soymilk (0.6 mg CAE/g) and tempeh day 2 (0.75 mg CAE/g) compared to natto, yogurt, tofu, and raw soybean (0.3-0.45 mg CAE/g) except soymilk slurry (0.89 mg CAE/g). Both types of cooked soymilk contained the highest (p<0.05) CTC. Fermented products, including natto, tempeh and yogurt contained higher (p <0.05) CTC compared to raw soybean. Among all products, pressed tofu had the lowest CTC, which was similar to that in soybean. Flavanol gallates has been reported to be converted into flavanols by esterase and tannase during fermentation by microorganisms [35].
Because of the complex systems of the range of soy foods, three antioxidant probes with different reaction mechanisms (single electron transfer for FRAP and DPPH assays, and hydrogen atom transfer for the ORAC assay) were used for antioxidant activity analyses [36]. Fermented products showed significantly higher DPPH activity (1.5-3.6 mg GAE/g) and FRAP activity (0.5-1.8 mg GAE/g) compared to soymilk, tofu and raw soybean (0.3-2.3 mg GAE/g). Among all soy products, pressed tofu was the lowest in DPPH radical scavenging and FRAP activities. ORAC values of natto day 2 and cooked soymilks (both traditional and cooked with okara) were significantly higher than that of tofu, natto day 4 and 6, tempeh, yogurt and raw soybean. Pressed tofu also had the lowest ORAC value. Compared to the common aging period of 2-3d [15], a longer aging period (6d) led to the decreases in phenolic content (TPC, TFC, CTC) and antioxidant activities in either natto or tempeh. After 2 days of fermentation with R. oligosporus, McCue and others [18] reported a decrease in soluble phenolic content by one-third that of the initial level. Compared to all processed products, pressed tofu, a non-fermented soy food with removed whey had the lowest phenolic contents and antioxidant capacity.
Processing conditions such as soaking, grinding, heat treatment, filtration, coagulation, solid-fermentation, and submerged fermentation showed variations in the phenolic content and antioxidant activities similar to other reports [12, 15, 27]. In general, phenolic compounds are responsible for nearly all antioxidant activities in soybean [6] and are highly correlated with each other [9, 27]. The antioxidant activities of polyphenols are due to 3’-4’ dihydroxy group in their B ring and the galolyl ester in the C ring of flavonoids [16]. Our results indicated that natto-2 day with the highest TPC was associated with the highest ORAC, and tempeh’s high TFC was associated with the high DPPH and FRAP, while cooked soymilks’ high CTC was associated with their high ORAC values. Our results substantiated that that variations in antioxidant activities of soybean and soy products may depend on both quantitative and qualitative compositions of antioxidants [37]. Bacillus subtilis (used for natto making) is capable of producing cellulase and β-glucanase [16] and Rhizopus oryzae (used for tempeh making) produces β-glucosidase, tannase and pectinase capable of breaking the cell wall matrix, resulting in the hydrolysis of ester bonds, which link phenolic compounds to cell wall and also in the oxidative degradation of lignin [17]. Similar to our results, B. subtilis has been reported to increase TPC in cheonggukjang and thua nua by 86% and 89%, respectively [24]. Results indicated that solid-state fermentation in natto yielded higher total TPC and CTC compared to Lactobacillus fermentation in soy yogurt [19]. The differences among different effects of fermentations may be due to the ability of the microorganisms to metabolize the phytochemicals, cellular localization of the β-glucosidase produced, culture conditions (semi-liquid or solid culture) or fermentation time. Bacterial produced β-glucosidases can be bound to cell wall or secreted into periplasmic space, while fungi synthesize cell-bound β-glucosidases in liquid conditions and secretes the enzymes in the environment under solid conditions [38, 39]. Georgetti and others [40] reported a higher activity of β-glucosidase on solid-state fermentation of soybean by Aspergillus species than during soymilk fermentation with Lactobacillus strains. An antioxidant activity is a reflection of an array of compounds rather than just total phenolic content [16]. To gain more insight of the relationship between chemical compositions and antioxidant capacity, further experiments were performed to identify and quantify individual phenolic acids and isoflavones in the soy foods.
Effect of processing on individual and total phenolic acid composition
In soybean, isoflavones are the major phenolic compounds, followed by phenolic acids and flavonoids [33]. Anthocyanin is present only in black soybean. The most important biological function of phenolic acids in plant cells is antioxidant abilities to protect against damage under stress-induced conditions. The free acids are scarcely deposited in high levels in the vacuoles of plant cells, while most are linked to cell wall structure (cellulose, hemicellulose, lignin, pectin and protein) through covalent bonds [17]. Fermentation [17, 41], thermal treatment [9, 15] and germination [12] have been reported to influence the yield and profiles of phenolic acids. However, to the best of our knowledge, a systematic comprehensive comparison across different processing of soy products is lacking. Table 1a and 1b present the free phenolic acid composition, and Table 2a and 2b present the total phenolic acid composition (free + conjugated) in soybean products. Fifteen individual phenolic acids were identified and quantified. The profile of phenolic acids differed in each processed product.
The free subtotal benzoic acids were higher (p <0.05) in natto and tempeh compared to tofu, soymilk and raw soybean (Table 1a) whereas natto and yogurt had higher (p <0.05) total free phenolic acid content than all other products (Tables 1a and 1b). The predominant phenolic acids in raw soybean were 2,3,4 trihydroxybenzoic (TBA), syringic acid among benzoics; p-coumaric + syringaldehyde (PCA+SD), m-coumaric, and o-coumaric acid among cinnamics. Tofu as a soy food category had the lowest total phenolic acids (Table 1b). The predominant acids in natto were TBA, protocatechuic and syringic, while gallic, TBA and syringic acids were the major phenolic acids in tempeh. Soymilk and yogurt were similar in phenolic acid profile including syringic, vanillic, PCA+SD, sinapic, and o-coumaric as the principal phenolic acids. Results indicated that Bacillus subtilis natto and Rhizopus oryzae released benzoic group, while lactic acid bacteria in yogurt led to increase in free cinnamic acid derivatives during fermentation. Dueñas and others [42] observed higher hydroxybenzoics (40.7-58.7 µg/g) than hydroxycinnamics (3.6-6.4 µg/g) during solid-state fermentation of soybean flour using Rhizopus oryzae and Bacillus subtilis similar to our results. Cho and others [35] observed significant increase in phenolic acids especially the benzoic derivatives, including gallic acid during fermentation by B. subtilis. Free benzoic acids in natto and tempeh might have contributed to higher antioxidant activity than cinnamic acids in yogurt. The antioxidant activity of phenolic acids depends on the number and orientation of the hydroxyl groups relative to the electron-withdrawing CO2H, CH2CO2H, or (CH)2CO2CH functional groups. The aromatic OH is a critical determinant of hydrogen donation and free radical scavenging by phenolic compounds [16, 43]. Similar to our result, Torino and others [41] reported that the release of bound phenolic acids into their free forms during fermentation improves the health-linked functionality in lentils. Storage/aging showed a decrease in free total phenolic acid composition in natto and tempeh. Chung and others [15] also observed 30-40% decrease in phenolic content during aging of ChungGukjang (soy paste) between 3d and 6d at 20°C.
Tables 2a and 2b show the total phenolic acid content (free + conjugated) in raw soybean (348 µg/g) was 14-91% higher (p <0.05) than all processed products, followed by yogurt (298-247 µg/g), natto (244-270 µg/g), cooked soymilk (219-253 µg/g), tempeh (89-127 µg/g) and tofu (31-50 µg/g). The loss of phenolic acids during food processing, such as steaming and boiling of soybeans (9), cooking of sprouts [12], soymilk processing (10), tempeh making [20] and natto making [15] have been reported. Phenolic acid compositions explained the high antioxidant activities in natto, cooked soymilk and yogurt. However, results indicated that compounds other than phenolic acids might also have contributed to the higher DPPH and FRAP values in tempeh and yoghurt since their ORAC (using a different antioxidant assay mechanism from DPPH and FRAP) values were lower than the original soybean (Figure 1). Lactic acid bacteria and Bacillus spp. had been evaluated for their potential to release phenolic acids as well as flavonoids from soybean [15, 35]. Soybean seeds fermented with Bacillus subtilis for three days yielded an increase in chlorogenic acid and naringin [15].
Effect of processing on composition of isoflavones
Tables 3a and 3b show the composition of isoflavones in the original raw soybean, soymilk, tofu, natto, tempeh and yogurt. The results were expressed in two ways similar to our previous study [12]: (1) Individual isoflavone contents of all isoflavones were calculated using response factors as described by AOAC method 2008.03 [34].
(2) Subtotal isoflavone contents of aglycones (aglycone equivalents) for each of the three types of isoflavones were calculated by converting the malonylglucosides, acetylglucosides, and β-glucosides weight into the aglycone weight using the respective molecular weight factors prior to summation. Total isoflavone contents were the sum of the adjusted sums of total genistein + total diadzein + total glycitein.
Results showed that the total isoflavone content in tempeh(s) (1728.3-1834.8 mg/g), natto(s) (1552.2-1887.0 mg/g) and pressed tofu (1677.1 mg/g), was significantly lower (p<0.05) than original soybean seed (2180.2 mg/g), cooked soymilk(s) (2100.4 -2214.4 mg/g), filled tofu (2160.8 mg/g), and yogurt(s) (2066.7-2263.0 mg/g). There were no statistical differences in the total isoflavone content, between the raw soybean, raw soymilk, filled tofu and most yogurt(s). Cooked soymilks were lower than raw soymilk, thus different from that reported by [44] who reported no changes in total isoflavone content in soymilks treated at 95°C and those treated with additional heating at 75°C. The discrepancy might be due to duration of cooking. Our traditional soymilks were boiled for 20 min to destroy trypsin inhibitors and lipoxygenases [45]. Filled tofu had higher total isoflavones than pressed tofu since filled tofu had not been pressed to release whey and therefore, retained the isoflavones in the whey fractions of the tofu making made by pressed method [15, 46]. Coagulation caused losses in total isoflavones during tofu making [7]. Tempeh might have lost most isoflavones during boiling or steaming [20] as the dehulling step of tempeh making has been reported to lose only 1.4% of isoflavones [46], and soybean hulls contain little amounts of isoflavones [47]. Cho and others (35) observed 64% decrease in total isoflavone content during 60 h fermentation, while Shin and others [24] reported 15% decrease in total isoflavone content during 48 h fermentation of cheonggukjang mostly due to the conversion of glycosides into aglycones. The high aglycones in tempeh may have significance in human nutrition as it has been recognized that isoflavone aglycones are absorbed faster [48] and in higher amounts than glucosides in humans [49]. Thus tempeh contained large amount of bioavailable phytoestrogens.
Different processing techniques significantly affected the profile of isoflavones in soybean products. Tempeh contained the highest amount of aglycones, including diadzein (ranging 430.1-451.2 mg/g for all days), genistein (980.8-1049.3 mg/g) and glycitein (53.5-54.6 mg/g), followed by raw or cooked soymilk (198.1- 412.0 mg/g; 200.5-259.3 mg/g; 27.9-52.6 mg/g, respectively, for the two types of soymilk), natto day 2 (149.8 mg/g, 130.4 mg/g , 16.0 mg/g, respectively ), filled tofu (127.2 mg/g, 177.1 mg/g, 18.3 mg/g, respectively), and pressed tofu (108.1 mg/g , 167.5 mg/g , 15.2 mg/, respectively). Raw soybean contained 70.4% malonyl glucosides, 22.3%, β-glucosides and 5.6% aglycones of the total isoflavones. Aglycones were the predominant isoflavone forms in tempeh, and it represented almost 74.6% of the total isoflavones followed by malonylglucosides (16.1-17.1%). Aglycones, β-glucosides and malonylglucosides represented 16.1-18.3%, 36.9-38.2% and 38.8-41.7%, respectively, of total isoflavones found in traditional cooked soymilk and soymilk cooked with okara. Natto day 2 was predominant in β-glucosides (87.8%) and malonylglucosides (8.4%). Tofu(s) and natto at day 4 and 6 were predominant in malonylglucosides (̴almost 58.9% and 59.3%, respectively) and β-glucoside forms (almost 29.4% and 30.6%, respectively). About 89.7% of isoflavones were in the β-glucoside forms in soy-yogurt. Pressed and filled tofu contained higher (p <0.05) amounts of MDin and MGin and lower (p <0.05) amount of Din, Gin, Dein, Gein, Glein and total isoflavones in comparison to both types of cooked soymilk(s).
In comparison to the original soybean seed, the combination of the soymilk processing steps of soaking, grinding and heating increased the aglycone content by 12 to 14 (p <0.05) fold in cooked soymilk(s), much higher than our report earlier on black soybean milk [8]. Different degrees of increases in aglycones also has been reported by our group and other researchers [7, 10, 44]. Soymilk cooked prior to okara separation exhibited higher (p<0.05) content of MGin, ADin, AGin, and lower (p<0.05) content of aglycones compared to traditional cooked soymilk, with no changes in the total isoflavone content. Aging of natto caused conversion of β-glucosides to malonylglucosides [50]. Toda and others [26] analyzed 50 commercial soy product samples and reported that the isoflavone profile of raw soybean and tofu were similar, containing 73% and 63% of total isoflavones in the forms of malonylglucosides. Commercial natto and soymilk were clustered together as high heat-treated products with respect to the profile of isoflavones. Similar to our results, reports have shown deglycosylation of isoflavone glycosides with fermentation by Bacillus subtilis [51], Rhizopus oligosporus [51] and lactic acid bacteria [52] through their β-glucosidase activities.
Aging/storage of natto drastically decreased β-glucosides and aglycones from day 2 to days 4 and 6, but increased malonylglucosides and acetylglucosides from day 2 to days 4 and 6. The total isoflavones increased at day 4 followed by a decrease on day 6. Aging/storage of tempeh caused gradual increases in aglycones, malonylglucosides, and acetylglucosides, but with decreases in β-glucosides till 6 days of storage, while no major changes were observed in yogurt for 8 days. Kuo and others [53] examined hydrolysis of isoflavone glycosides by B. subtilis natto in black soymilk, and found that the activity of β-glucosidase increased till 12 h and then decreased rapidly till 36 h. McCue and others [18] reported that R. oligosporus fermentation on soybean powder showed β-glucosidase activity that increased during mid to late stages of fermentation from 0-10 days and correlated with total phenolic content. However, in our study the increases in aglycones were not enough to change the antioxidant or phenolic content.
Correlations and Principal Component Analysis (PCA) of phenolic compounds and antioxidant activities of fermented and non-fermented soybean products
The linear correlation coefficients between 55 variables (TPC, TFC, CTC 36 free and conjugated phenolic variables explained in Tables 1 and 2, and 16 isoflavones in Table 3) and antioxidant activities in 18 fermented and non-fermented products are shown in Table 4. Twenty-nine variables were found to be significantly correlated with either DPPH, FRAP or ORAC in fermented products, and nineteen variables were correlated (p<0.05) with antioxidant activities in non-fermented products. In fermented products 14 variables, and in non-fermented products 10 variables were positively correlated (p <0.05) with ORAC. Total flavonoid content, free phenolic acids such as- protocatechuic acid, hydroxybenzoic acid, 2,3,4-trihydroxybenzoic acid, total genistic acid, vanillic acid, glycitin, malonyl glycitin, subtotal genistein, subtotal glycitein and total isoflavones were positively correlated with DPPH or FRAP. The negative correlations indicate conversion or degradation of phenolic compounds with respect to change in antioxidant activities. The results showed that phytochemicals play an essential role in the antioxidant activities of soybean products.
PCA was performed to visualize the main differences, cluster trends or outliers among soybean products as affected by processing techniques in terms of phytochemicals and antioxidant properties. Figure 2a-b shows the two-dimensional score plot (PC1 vs PC2 and PC1 vs. PC3) derived from computing 20 variables and 18 products (PC1, PC2, and PC3 explained 36.9%, 20.5%, and 17.7% of the total variance, respectively). In other words, 75.1% of the total variance in the 20 considered variables can be condensed into three new variables (PCs). Correspondingly, tempeh day 2, 4 and 6 were characterized with higher aglycones content explaining higher FRAP and DPPH value, yogurt day 0, 2, 4, 6, 8 were explained with higher phenolic acids and total isoflavones imparting to higher DPPH and FRAP values. Natto day 2 was characterized with exceptionally higher total phenolic content, condensed tannin content, free benzoics leading to higher ORAC.
Agglomerative hierarchical cluster analysis was conducted based on the distribution of 20 variables (Figure 3). Classifications were determined as follows: A, tempeh (day 2, 4, 6), pressed tofu and filled tofu; B, yogurt (day 0, 2, 4, 6, 8); and C, raw soybean, cooked soymilk(s) and natto (day 2, 4, 6). Toda and others (26) clustered 50 samples of commercial soybean foods into 6 clusters based on the distribution of isoflavones and described that each cluster was characterized by the effects of the heating method and temperature. Natto and cooked soymilk was clustered together based on high heat treatment compared to tofu in another cluster. Heat treatment and fermentation dependent on microorganism reflected the three clusters.
CONCLUSIONS
We have achieved our goal in comparing the relative enhancement or damage of phenolic compounds, isoflavones, and antioxidant properties as affected by several processing methods that are commonly used for making traditional soy foods. Such comparative information was possible and novel since the wide variation due to raw material differences as presented in most previous literature has been eliminated by using one single soybean cultivar. For consumer health promotion as relevant to the compositions of isoflavones, other phenolic substances, and the overall antioxidant capacity, different soy foods have different potentials as they contain different proportions of the compounds. In general, processing for fermented foods has enhanced more than that for non-fermented foods in terms of antioxidant properties. However, soymilks and filled tofu among non-fermented foods tested also retained most of the antioxidant factors in soybeans. Pressed tofu still retained substantial amount of phytoestrogen isoflavones and phenolic acids, even though it ranked the lowest among all soy foods tested.
Heat treatment and fermentation as a function of microorganism clustered all products into 3 groups 1. tempeh and tofu (s); 2. natto, cooked soymilk(s), and raw soybean; and 3. yogurt. Filled tofu contained higher phenolic content than pressed tofu, while no major difference was observed between traditional cooked soymilk and soymilk cooked with okara. Cooked soymilk (s) was characterized with high TFC, CTC and aglycones. Solid fermentation by B. subtilis released higher free and total phenolic acids while tempeh was characterized by highest content in aglycones compared to set-style yogurt with higher content of free cinnamic acids and isoflavone β-glucosides. Free benzoic acids and aglycones had higher antioxidant activities compared to cinnamic acids and β-glucosides. With the present scope of this study, tempeh (high DPPH and FRAP), natto (high ORAC) and cooked soymilk (high ORAC) can be preferred over other products.
However, further studies to demonstrate the advantages of the unique groups of phenolic compounds in particular soy foods are needed to elucidate information regarding their bioavailability and bioactivities in humans.
Acknowledgments
USDA-ARS SCA no. 58-6402-2729 and 58-6066-7081 contributed funding for this CRIS project number. MIS 081710. We thank Mr. Yan Zhang for his technical assistance on phenolic and antioxidant analyses. This work is a contribution of Mississippi Agricultural and Forestry Experiment Station, Mississippi State University.
REFERENCES
1. Tsukamoto Y, Ichise H, Kakuda H, Yamaguchi M. Intake of fermented soybean ( natto ) increases circulating vitamin K 2. (menaquinone-7) and γ-carboxylated osteocalcin concentration in normal individuals. Journal of Bone and Mineral Metabolism. 2000;18(4):216-22.
2. Liu J, Chang SKC, Wiesenborn D. Antioxidant Properties of Soybean Isoflavone Extract and Tofu in Vitro and in Vivo. Journal of Agricultural and Food Chemistry. 2005;53(6):2333-40.
3. Park D, Huang T, Frishman WH. Phytoestrogens as Cardioprotective Agents. Cardiology in Review. 2005;13(1):13-7.
4. Yao F-R, Sun C-W, Chang SKC. Morton Lentil Extract Attenuated Angiotensin II-Induced Cardiomyocyte Hypertrophy via Inhibition of Intracellular Reactive Oxygen Species Levels in Vitro. Journal of Agricultural and Food Chemistry. 2010;58(19):10382-8.
5. Valko M, Leibfritz D, Moncol J, Cronin MTD, Mazur M, Telser J. Free radicals and antioxidants in normal physiological functions and human disease. The International Journal of Biochemistry & Cell Biology. 2007;39(1):44-84.
6. Pratt DE. Natural Antioxidants of Soybeans and Other Oil-Seeds. Autoxidation in Food and Biological Systems: Springer US; 1980. p. 283-93.
7. Jackson CJC, Dini JP, Lavandier C, Rupasinghe HPV, Faulkner H, Poysa V, et al. Effects of processing on the content and composition of isoflavones during manufacturing of soy beverage and tofu. Process Biochemistry. 2002;37(10):1117-23.
8. Tan Y, Chang SKC, Zhang Y. Innovative Soaking and Grinding Methods and Cooking Affect the Retention of Isoflavones, Antioxidant and Antiproliferative Properties in Soymilk Prepared from Black Soybean. Journal of Food Science. 2016;81(4):H1016-H23.
9. Xu B, Chang SKC. Isoflavones, Flavan-3-ols, Phenolic Acids, Total Phenolic Profiles, and Antioxidant Capacities of Soy Milk As Affected by Ultrahigh-Temperature and Traditional Processing Methods. Journal of Agricultural and Food Chemistry. 2009;57(11):4706-17.
10. Zhang Y, Chang SKC, Liu Z. Isoflavone Profile in Soymilk as Affected by Soybean Variety, Grinding, and Heat-Processing Methods. Journal of Food Science. 2015;80(5):C983-C8.
11. Kim EH, Kim SH, Chung JI, Chi HY, Kim JA, Chung IM. Analysis of phenolic compounds and isoflavones in soybean seeds (Glycine max (L.) Merill) and sprouts grown under different conditions. European Food Research and Technology. 2005;222(1-2):201-8.
12. Kumari S, Chang SKC. Effect of Cooking on Isoflavones, Phenolic Acids, and Antioxidant Activity in Sprouts of Prosoy Soybean (Glycine max). Journal of Food Science. 2016;81(7):C1679-C91.
13. Xu B, Chang SKC. Reduction of antiproliferative capacities, cell-based antioxidant capacities and phytochemical contents of common beans and soybeans upon thermal processing. Food Chemistry. 2011;129(3):974-81.
14. Xu B, Chang SKC. Effect of soaking, boiling, and steaming on total phenolic contentand antioxidant activities of cool season food legumes. Food Chemistry. 2008;110(1):1-13.
15. Chung I-M, Seo S-H, Ahn J-K, Kim S-H. Effect of processing, fermentation, and aging treatment to content and profile of phenolic compounds in soybean seed, soy curd and soy paste. Food Chemistry. 2011;127(3):960-7.
16. Hur SJ, Lee SY, Kim Y-C, Choi I, Kim G-B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chemistry. 2014;160:346-56.
17. Huynh NT, Van Camp J, Smagghe G, Raes K. Improved release and metabolism of flavonoids by steered fermentation processes: a review. Int J Mol Sci. 2014;15(11):19369-88.
18. McCue P, Horii A, Shetty K. Solid-state bioconversion of phenolic antioxidants from defatted soybean powders by Rhizopus oligosporus: role of carbohydrate-cleaving enzymes. Journal of Food Biochemistry. 2003;27(6):501-14.
19. Martins S, Mussatto SI, Martínez-Avila G, Montañez-Saenz J, Aguilar CN, Teixeira JA. Bioactive phenolic compounds: Production and extraction by solid-state fermentation. A review. Biotechnology Advances. 2011;29(3):365-73.
20. Haron H, Raob N. Changes in macronutrient, total phenolic and anti-nutrient contents during preparation of tempeh. Nutrition & Food Sciences. 2014.
21. Hati S, Vij S, Singh BP, Mandal S. β -Glucosidase activity and bioconversion of isoflavones during fermentation of soymilk. Journal of the Science of Food and Agriculture. 2014;95(1):216-20.
22. Marazza JA, LeBlanc JG, de Giori GS, Garro MS. Soymilk fermented with Lactobacillus rhamnosus CRL981 ameliorates hyperglycemia, lipid profiles and increases antioxidant enzyme activities in diabetic mice. Journal of Functional Foods. 2013;5(4):1848-53.
23. Juan M-Y, Chou C-C. Enhancement of antioxidant activity, total phenolic and flavonoid content of black soybeans by solid state fermentation with Bacillus subtilis BCRC 14715. Food Microbiology. 2010;27(5):586-91.
24. Shin E-C, Lee JH, Hwang CE, Lee BW, Kim HT, Ko JM, et al. Enhancement of total phenolic and isoflavone-aglycone contents and antioxidant activities during Cheonggukjang fermentation of brown soybeans by the potential probiotic Bacillus subtilis CSY191. Food Science and Biotechnology. 2014;23(2):531-8.
25. Dajanta K, Apichartsrangkoon A, Chukeatirote E. Antioxidant Properties and Total Phenolics of Thua Nao (a Thai Fermented Soybean)as Affected by Bacillus-fermentation. Journal of Microbial & Biochemical Technology. 2011;03(04).
26. Toda T, Sakamoto A, Takayanagi T, Yokotsuka K. Changes in Isoflavone Compositions of Soybean Foods during Cooking Process. Food Science and Technology Research. 2000;6(4):314-9.
27. Xu B, Chang SKC. Total Phenolics, Phenolic Acids, Isoflavones, and Anthocyanins and Antioxidant Properties of Yellow and Black Soybeans As Affected by Thermal Processing. Journal of Agricultural and Food Chemistry. 2008;56(16):7165-75.
28. Handa CL, Zhang Y, Kumari S, Xu J, Ida EI, Chang SK. Comparative Study of Angiotensin I-Converting Enzyme (ACE) Inhibition of Soy Foods as Affected by Processing Methods and Protein Isolation. Processes. 2020;8(8):978.
29. Zhang Y, Guo S, Liu Z, Chang SKC. Off-Flavor Related Volatiles in Soymilk As Affected by Soybean Variety, Grinding, and Heat-Processing Methods. Journal of Agricultural and Food Chemistry. 2012;60(30):7457-62.
30. Meng S, Chang S, Gillen AM, Zhang Y. Protein and quality analyses of accessions from the USDA soybean germplasm collection for tofu production. Food Chemistry. 2016;213:31-9.
31. Donkor ON, Henriksson A, Vasiljevic T, Shah NP. Rheological Properties and Sensory Characteristics of Set-Type Soy Yogurt. Journal of Agricultural and Food Chemistry. 2007;55(24):9868-76.
32. Wang Y-C, Yu R-C, Chou C-C. Growth and survival of bifidobacteria and lactic acid bacteria during the fermentation and storage of cultured soymilk drinks. Food Microbiology. 2002;19(5):501-8.
33. Xu B, Chang SKC. Characterization of Phenolic Substances and Antioxidant Properties of Food Soybeans Grown in the North Dakota−Minnesota Region. Journal of Agricultural and Food Chemistry. 2008;56(19):9102-13.
34. Collison MW. Determination of total soy isoflavones in dietary supplements, supplement ingredients, and soy foods by high-performance liquid chromatography with ultraviolet detection: collaborative study. Journal of AOAC International. 2008;91(3):489-500.
35. Cho K, Hong S, Math R, Lee J, Kambiranda D, Kim J, et al. Biotransformation of phenolics (isoflavones, flavanols and phenolic acids) during the fermentation of cheonggukjang by Bacillus pumilus HY1. Food Chemistry. 2009;114(2):413-9.
36. Prior RL, Wu X, Schaich K. Standardized Methods for the Determination of Antioxidant Capacity and Phenolics in Foods and Dietary Supplements. Journal of Agricultural and Food Chemistry. 2005;53(10):4290-302.
37. Lee SJ, Yan W, Ahn JK, Chung IM. Effects of year, site, genotype and their interactions on various soybean isoflavones. Field Crops Research. 2003;81(2-3):181-92.
38. Iwashita K, Shimoi H, Ito K. Extracellular soluble polysaccharide (ESP) from Aspergillus kawachii improves the stability of extracellular β-gluocosidases (EX-1 and EX-2) and is involved in their localization. Journal of Bioscience and Bioengineering. 2001;91(2):134-40.
39. Pyo Y-H, Lee T-C, Lee Y-C. Enrichment of bioactive isoflavones in soymilk fermented with β-glucosidase-producing lactic acid bacteria. Food Research International. 2005;38(5):551-9.
40. Georgetti SR, Vicentini FTMC, Yokoyama CY, Borin MF, Spadaro ACC, Fonseca MJV. Enhancedin vitroandin vivoantioxidant activity and mobilization of free phenolic compounds of soybean flour fermented with different β-glucosidase-producing fungi. Journal of Applied Microbiology. 2009;106(2):459-66.
41. Torino MI, Limón RI, Martínez-Villaluenga C, Mäkinen S, Pihlanto A, Vidal-Valverde C, et al. Antioxidant and antihypertensive properties of liquid and solid state fermented lentils. Food Chemistry. 2013;136(2):1030-7.
42. Dueñas M, Fernández D, Hernández T, Estrella I, Muñoz R. Bioactive phenolic compounds of cowpeas (Vigna sinensisL). Modifications by fermentation with natural microflora and withLactobacillus plantarumATCC 14917. Journal of the Science of Food and Agriculture. 2004;85(2):297-304.
43. Ng TB, Liu F, Wang ZT. Antioxidative activity of natural products from plants. Life Sciences. 2000;66(8):709-23.
44. Jung S, Murphy PA, Sala I. Isoflavone profiles of soymilk as affected by high-pressure treatments of soymilk and soybeans. Food Chemistry. 2008;111(3):592-8.
45. Yuan S, Chang SK, Liu Z, Xu B. Elimination of trypsin inhibitor activity and beany flavor in soy milk by consecutive blanching and ultrahigh-temperature (UHT) processing. Journal of agricultural and food chemistry. 2008;56(17):7957-63.
46. Wang H-J, Murphy PA. Mass Balance Study of Isoflavones during Soybean Processing. Journal of Agricultural and Food Chemistry. 1996;44(8):2377-83.
47. Kudou S, Tsuizaki I, Uchida T, Okubo K. Purification and some properties of soybean saponin hydrolase from Aspergillus oryzae KO-2. Agricultural and Biological Chemistry. 1991;55(1):31-6.
48. Setchell KDR, Brown NM, Zimmer-Nechemias L, Brashear WT, Wolfe BE, Kirschner AS, et al. Evidence for lack of absorption of soy isoflavone glycosides in humans, supporting the crucial role of intestinal metabolism for bioavailability. The American Journal of Clinical Nutrition. 2002;76(2):447-53.
49. Izumi T, Piskula MK, Osawa S, Obata A, Tobe K, Saito M, et al. Soy Isoflavone Aglycones Are Absorbed Faster and in Higher Amounts than Their Glucosides in Humans. The Journal of Nutrition. 2000;130(7):1695-9.
50. Rostagno MA, Villares A, Guillamón E, García-Lafuente A, Martínez JA. Sample preparation for the analysis of isoflavones from soybeans and soy foods. Journal of Chromatography A. 2009;1216(1):2-29.
51. Cheng K-C, Wu J-Y, Lin J-T, Liu W-H. Enhancements of isoflavone aglycones, total phenolic content, and antioxidant activity of black soybean by solid-state fermentation with Rhizopus spp. European Food Research and Technology. 2013;236(6):1107-13.
52. Choi H-J, Cheigh C-I, Kim S-B, Lee J-C, Lee D-W, Choi S-W, et al. Weissella kimchii sp. nov., a novel lactic acid bacterium from kimchi. International Journal of Systematic and Evolutionary Microbiology. 2002;52(2):507-11.
53. Kuo L-C, Cheng W-Y, Wu R-Y, Huang C-J, Lee K-T. Hydrolysis of black soybean isoflavone glycosides by Bacillus subtilis natto. Applied Microbiology and Biotechnology. 2006;73(2):314-20.
PEER REVIEW
Not commissioned. Externally peer reviewed.
FIGURES
Figure 1. Total phenolic compositions and antioxidant activities of fermented and non-fermented soybean products Bars are expressed as the mean ± standard deviation (n=2) on dry weight basis; values marked with different letter above the bars are significantly different (p <0.05). RTSmilk, raw traditional soymilk; CTSmilk, cooked traditional soymilk; RSmilk slurry, raw soymilk slurry; CSmilkOF, cooked soymilk with okra and filtered.
Figure 2. Scatterplot of the PCA analysis (a) representing 20 variables and (b) representing 18 soybean products. Principal component numbers 1, 2 and 3 (PC1, PC2, PC3) were computed applying 20 variables containing phenolics content, free phenolics, conjugated phenolics, isoflavones and antioxidant properties, and explained 36.9%, 20.5% and 17.5%, respectively, of the total variance in the data. Variables: F-Ben, Free phenolic acid analysis- subtotal benzoics; F-Cin, Free phenolic acid- subtotal cinnamics; F-TP, Free phenolic acid- total phenolics content; C-Ben, Conjugated phenolic acid analysis- subtotal benzoics; C-Cin, Conjugated phenolic acid analysis- subtotal cinnamics; C-TP, Conjugated phenolic acid analysis- total phenolics content; β- glucosides, total of Din, Gin and Gly; M-glucosides, total of MDin, MGin and MGly; A-glucosides, total of ADin, AGin and AGly; Aglycones, total of Dein, Gein and Glein; T-Dein, TGein, T-Glein is subtotal individuals = moles of isoflavone × molecular weight of aglycone form isoflavone. RTSmilk, raw traditional soymilk; CTSmilk, cooked traditional soymilk; RSmilk slurry, raw soymilk slurry; CSmilkOF, cooked soymilk with okra and filtered.
Figure 3. Dendrogram of raw soybean and processed products connected on the basis of the distribution of 20 variables. Variables: F-Ben, Free phenolic acid analysis- subtotal benzoics; F-Cin, Free phenolic acid- subtotal cinnamics; F-TP, Free phenolic acid- total phenolics content; C-Ben, Conjugated phenolic acid analysis- subtotal benzoics; C-Cin, Conjugated phenolic acid analysis- subtotal cinnamics; C-TP, Conjugated phenolic acid analysis- total phenolics content; β- glucosides, total of Din, Gin and Gly; M-glucosides, total of MDin, MGin and MGly; A-glucosides, total of ADin, AGin and AGly; Aglycones, total of Dein, Gein and Glein; T-Dein, TGein, T-Glein is subtotal individuals = moles of isoflavone × molecular weight of aglycone form isoflavone. RTSmilk, raw traditional soymilk; CTSmilk, cooked traditional soymilk; RSmilk slurry, raw soymilk slurry; CSmilkOF, cooked soymilk with okra and filtered; TempD2; Tempeh day 2; Yogurt D0, yogurt day 0; NattoD2, natto day 2.
TABLES
Table 1a. Free phenolic acid compositions (µg/g) in fermented and non-fermented soybean products a
|
Samples |
Individual benzoic acid derivative |
Subtotal benzoics |
||||||||
|
|
GA |
PA |
TBA |
PCD |
HBA |
GNA |
SA |
VA |
VN |
|
|
Soybean |
16.8 ± 0.2 d |
9.7 ± 0.2 cde |
8 ± 0.6 fghi |
2.2 ± 1.2 fg |
3.4 ± 0.4 c |
4.1 ± 3.5 de |
16.6 ± 1.1 defgh |
21.9 ± 4.5 a |
2.2 ± 1 gh |
84.9 ± 8.7 h |
|
RTSmilk |
11.5 ± 0.8 d |
8.1 ± 0.5 de |
6.8 ± 0.2 ghi |
4.1 ± 0.1 e |
4.1 ± 0.1 b |
10.2 ± 0.3 a |
23.8 ± 0 bc |
14.7 ± 0.3 bcd |
3.3 ± 0.2 ef |
86.6 ± 0.4 fg |
|
CTSmilk |
13.2 ± 1.1 d |
10.2 ± 0.2 cde |
8.6 ± 0.2 fghi |
6.9 ± 0.3 c |
3.2 ± 0.2 cd |
8.1 ± 0.3 ab |
24.1 ± 2.8 bc |
15.7 ± 0.9 bc |
5.3 ± 0.4 abc |
95.3 ± 5.5 fe |
|
RSmilk slurry |
1.8 ± 0.1 f |
7.1 ± 0.5 e |
5.6 ± 0.4 hi |
0.6 ± 0.2 cde |
2 ± 0.1 def |
5.3 ± 0.2 bcd |
13.9 ± 1 fgh |
8.9 ± 1.8 h |
3.5 ± 0.3 e |
48.7 ± 3.7 i |
|
CSmilkOF |
14.5 ± 0.6 d |
10.3 ± 0.1 cde |
9.3 ± 0.2 fgh |
5.6 ± 0.4 gh |
2.8 ± 0.2 ih |
6.7 ± 1.2 bcd |
23.2 ± 4.8 bcd |
14.8 ± 0.6 bcd |
4.8 ± 0.2 bcd |
92 ± 5.1 fe |
|
Pressed tofu |
4.2 ± 0.1 ef |
3.6 ± 0.2 e |
3.7 ± 0 i |
0.2 ± 0 h |
nd |
nd |
0.9 ± 0.0 j |
4.8 ± 0.1 i |
0.8 ± 0.0 i |
18.2 ± 0.2 j |
|
Filled tofu |
6.4 ± 0.1 e |
3.7 ± 0.1 e |
3.8 ± 0 i |
0.3 ± 0 h |
nd |
nd |
15.5 ± 0.1 fgh |
16 ± 3.8 bc |
0.8 ± 0.0 i |
46.5 ± 0.2 i |
|
Natto D2 |
28.2 ± 4.7 a |
30 ± 6 a |
12.6 ± 1.7 f |
33.7 ± 2.6 a |
6 ± 0.4 a |
6.8 ± 0.3 bcd |
42.1 ± 3.4 a |
14.3 ± 0.6 bcde |
5.9 ± 0.1 a |
179.6 ± 8.4 a |
|
Natto D4 |
22 ± 3.3 c |
18.9 ± 1.5 bc |
12.8 ± 0.1 f |
16.8 ± 0.9 b |
4.1 ± 0.2 b |
4.4 ± 0.3 cde |
23.1 ± 3.0 bcde |
12.4 ± 1.3 defg |
5.5 ± 0 abc |
119.9 ± 0.2 bc |
|
Natto D6 |
24.7 ± 1.6 acb |
18.8 ± 4.5 bc |
11.5 ± 0.4 fg |
16.9 ± 0.6 b |
4.4 ± 0.5 b |
5.9 ± 0.3 bcd |
25.4 ± 3.7 b |
13.5 ± 0.2 cdef |
5.3 ± 0.4 abc |
126.4 ± 0.4 b |
|
Tempeh D2 |
28.5 ± 2.2 a |
26.3 ± 2.5 b |
28.8 ± 2.2 c |
2.4 ± 0.2 f |
2.1 ± 0.1 ghi |
1.9 ± 0.1 fe |
17.5 ± 1.6 cdefg |
15.2 ± 0.9 bcd |
2.5 ± 0.2 fg |
125.1 ± 9.7 bc |
|
Tempeh D4 |
24.1 ± 0.3 cb |
20.3 ± 1.7 b |
25.5 ± 1.5 cde |
1.5 ± 0.1 fgh |
1.8 ± 0 i |
2 ± 0.2 fe |
12.2 ± 0.3 gh |
14.4 ± 0.4 bcde |
1.4 ± 0.3 hi |
103.2 ± 3.4 de |
|
Tempeh D6 |
26.4 ± 3.8 ab |
17.3 ± 0.7 bcd |
28.1 ± 0.8 cd |
1.2 ± 0.2 fgh |
1.9 ± 0.1 hi |
2.2 ± 0.2 fe |
10.1 ± 0.8 hi |
17 ± 3.6 b |
1 ± 0.3 i |
105.3 ± 8 de |
|
Yogurt D0 |
3.1 ± 0.2 ef |
5.8 ± 0.6 e |
23.2 ± 0.9 e |
4.4 ± 0.1 e |
2.5 ± 0.2 efg |
6.3 ± 0.1 bcd |
13.8 ± 0.9 fgh |
11.3 ± 0.3 efgh |
4.6 ± 0.3 cd |
75.1 ± 2.3 g |
|
Yogurt D2 |
3.9 ± 1 ef |
6.8 ± 0.5 e |
28.3 ± 3.5 cd |
5.3 ± 0.1 cde |
2.8 ± 0.1 def |
6.3 ± 0.3 bcd |
16.3 ± 1.5 defgh |
13.4 ± 0.5 cdef |
4 ± 0.1 de |
87.1 ± 4.8 fg |
|
Yogurt D4 |
4.4 ± 0.3 ef |
6.6 ± 0.2 e |
42 ± 8.2 a |
6.4 ± 0.3 cd |
3 ± 0.2 def |
6.7 ± 0.6 bcd |
20.4 ± 0.8 bcdef |
16.8 ± 1.7 b |
4.7 ± 0.2 cd |
111 ± 10.8 dc |
|
Yogurt D6 |
4 ± 0.4 ef |
6 ± 0.3 e |
33.6 ± 0.5 b |
5 ± 0.4 de |
2.7 ± 0 def |
7.3 ± 0.9 bcd |
16.1 ± 1.7 efgh |
10.4 ± 0.1 fgh |
5.8 ± 0.9 ab |
90.9 ± 2.2 fe |
|
Yogurt D8 |
3.2 ± 0.2 ef |
4.5 ± 0.1 e |
40.8 ± 0.2 a |
4.7 ± 0.2 de |
2.4 ± 0 fgh |
6.1 ± 0.4 bcd |
17.7 ± 3.3 cdefg |
9.6 ± 0.2 gh |
4.6 ± 0.2 cd |
93.5 ± 3.4 fe |
|
Samples |
Individual cinnamic acid derivative |
subtotal cinnamics |
total free phenolic acids |
|||||
|
|
CLA |
PCA+SD |
MCA |
FA |
SPA |
OCA |
||
|
Soybean |
6.3 ± 0.7 ab |
12.4 ± 2.1 b |
6.7 ± 1 fg |
9.3 ± 1.3 ab |
7.9 ± 0.7 efg |
13.1 ± 0.6 g |
55.8 ± 5.5 e |
140.5 ± 6.3 h |
|
RTSmilk |
6.8 ± 0.2 ab |
9.6 ± 0.5 cdef |
22.7 ± 0.5 a |
6.9 ± 2.8 cb |
8.8 ± 0.1 def |
15.1 ± 0.4 fg |
70.6 ± 3.6 cd |
157.3 ± 3.2 fgh |
|
CTSmilk |
6.9 ± 0.3 ab |
9.5 ± 0 cdef |
8.4 ± 1.2 ef |
2.6 ± 2 e |
10.8 ± 0.2 d |
20.4 ± 0.8 e |
59.2 ± 1.1 e |
154.5 ± 4.4 gh |
|
RSmilk slurry |
3.4 ± 0.3 cde |
8.6 ± 0.1 defg |
3.6 ± 0.7 hi |
2.2 ± 1.4 e |
5.6 ± 0.3 gh |
7.2 ± 1.1 h |
30.8 ± 4 g |
79.5 ± 7.7 j |
|
CSmilkOF |
6.2 ± 0.2 ab |
9.4 ± 0.2 cdef |
8.3 ± 0.8 efg |
2.5 ± 1.9 e |
10.5 ± 0.6 de |
18.8 ± 2.4 ef |
56.3 ± 2.8 e |
148.2 ± 2.3 gh |
|
Pressed tofu |
1.3 ± 0.0 e |
6.9 ± 1.4 gh |
1.6 ± 0.1 i |
1.1 ± 0 e |
5.3 ± 0.3 gh |
nd |
16.2 ± 0.4 h |
34.5 ± 0.6 k |
|
Filled tofu |
6.8 ± 4.9 ab |
8.2 ± 0.2 defg |
11.8 ± 0 cd |
3.1 ± 0 ed |
15.7 ± 0.1 c |
36.4 ± 0 c |
82.0 ± 4.9 bc |
128.5 ± 5.1 hi |
|
Natto D2 |
8.6 ± 0.4 a |
10.1 ± 0.7 bcd |
5.6 ± 2.3 gh |
5.6 ± 0.2 cd |
8.1 ± 1.9 defg |
13.1 ± 0.2 g |
51.1 ± 5.2 e |
230.7 ± 6.1 a |
|
Natto D4 |
5.5 ± 0.4 bc |
9.4 ± 0.1 cdef |
5.7 ± 1.1 gh |
4 ± 0.1 ed |
6.8 ± 0.2 fg |
12.6 ± 3.4 g |
44.4 ± 4.9 f |
164.3 ± 5.1 gfe |
|
Natto D6 |
6.2 ± 0.2 ab |
10 ± 0.6 cdef |
6.6 ± 0.1 fg |
3.7 ± 0.4 ed |
6.2 ± 1 fgh |
11.3 ± 0.9 g |
44.3 ± 1.1 f |
170.7 ± 1.5 dfe |
|
Tempeh D2 |
2 ± 0 de |
8.6 ± 0.7 defg |
2.1 ± 0.1 i |
1.5 ± 0 e |
3.9 ± 0 h |
0.9 ± 0.2 i |
18.9 ± 0.9 h |
144 ± 8.8 gh |
|
Tempeh D4 |
1.9 ± 0 de |
7.7 ± 0.3 efgh |
1.9 ± 0.1 i |
1.5 ± 0.1 e |
3.4 ± 0 h |
1.2 ± 0.1 i |
18.1 ± 0.1 h |
121.3 ± 3.5 i |
|
Tempeh D6 |
1.8 ± 0.1 de |
7.4 ± 0.2 fgh |
2.3 ± 0.1 i |
1.5 ± 0 e |
3.3 ± 0 h |
1.1 ± 0.1 i |
18.3 ± 0.3 h |
123.3 ± 7.7 i |
|
Yogurt D0 |
5.1 ± 0.4 bc |
11.5 ± 1.8 bc |
10.3 ± 1.1 de |
7.7 ± 0.4 cb |
15.9 ± 1 c |
32.7 ± 4.2 d |
89.1 ± 8.2 b |
158.2 ± 10.4 fgh |
|
Yogurt D2 |
5.2 ± 0.7 bc |
9.8 ± 1.4 cdef |
11.7 ± 1.4 cd |
8.7 ± 1.6 ab |
16.1 ± 1.1 c |
37.5 ± 4.6 c |
88.9 ± 10.8 b |
175.8 ± 15.6 de |
|
Yogurt D4 |
6.1 ± 0.3 abc |
11.5 ± 1.9 bc |
12.9 ± 2.8 c |
10.9 ± 0.7 a |
19.3 ± 0.4 b |
47.4 ± 1.9 a |
108.1 ± 4.2 a |
219.1 ± 6.7 b |
|
Yogurt D6 |
5.6 ± 0.4 bc |
11.5 ± 0.2 bc |
13.2 ± 0.1 c |
8.3 ± 1.7 ab |
9.6 ± 2.4 b |
42.5 ± 0.2 b |
91 ± 0.1 b |
181.9 ± 2.1 d |
|
Yogurt D8 |
4.7 ± 0.4 bc |
14.8 ± 0.4 a |
16.2 ± 1.5 b |
4 ± 0.4 ed |
27 ± 3.6 a |
39.5 ± 1.7 bc |
106.1 ± 2.2 a |
199.7 ± 1.2 c |
Table 1b. Free phenolic acid compositions (µg/g) in fermented and non-fermented soybean products a (continued from 1a)
a Data are expressed as means of triplicate experiments on dry weight basis. Values marked by the same letter within each column are not significantly different (p > 0.05). Phenolic acids: GA, gallic acid; PA, protocatechuic acid; TBA, 2,3,4-trihydroxybenzoic acid; PCD, protocatechualdehyde; HBA, p-hydroxybenzoic acid; GNA, gentistic acid; SA, syringic acid; VA, vanillic acid; VN, vanillin; CLA, chlorogenic acid; PCA + SD, p-coumaric acid + syringaldehyde; MCA + FA , m-coumaric acid + ferullic acid; SPA, sinapic acid; OCA, o-coumaric acid. ND, not detectable; RTSmilk, raw traditional soymilk; CTSmilk, cooked traditional soymilk; RSmilk slurry, raw soymilk slurry; CSmilkOF, cooked soymilk with okra and filtered.
Table 2a. Total phenolic acid compositions (µg/g) in fermented and non-fermented soybean products a
|
Samples |
Individual benzoic acid derivative |
Subtotal benzoics |
||||||||
|
|
GA |
PA |
TBA |
PCD |
HBA |
GNA |
SA |
VA |
VN |
|
|
Soybean |
12.9 ± 0.6 bc |
6.4 ± 0 cde |
85.9 ± 4.1 a |
2 ± 0.2 c |
3.4 ± 0.2 c |
2.4 ± 0.1 abcd |
68.3 ± 10 a |
16.9 ± 0.8 b |
1.6 ± 0.2 b |
199.9 ± 12.8 a |
|
RTSmilk |
5.3 ± 0.5 hi |
5.2 ± 0.3 ef |
34 ± 0.1 de |
1.5 ± 0.1 c |
3.4 ± 0.2 c |
1.5 ± 0.3 de |
52.5 ± 0.2 d |
12.9 ± 0.3 b |
1.6 ± 0.1 b |
117.9 ± 0.7 def |
|
CTSmilk |
10.9 ± 2 cde |
5.8 ± 0.4 de |
7.5 ± 0.5 h |
1.1 ± 0.1 c |
4.6 ± 0.1 b |
2.5 ± 0.2 abc |
61 ± 6.8 abcd |
15.5 ± 0.9 b |
3.9 ± 3.7 b |
112.9 ± 5 ef |
|
RSmilk slurry |
6.2 ± 0.1 hi |
5.9 ± 0.1 de |
9 ± 0.3 h |
0.8 ± 0 c |
2.1 ± 0.1 def |
1.5 ± 0.3 e |
55.5 ± 0.9 cd |
13.3 ± 0 b |
1.7 ± 0 a |
95.9 ± 1 fg |
|
CSmilkOF |
10 ± 0.3 def |
5.5 ± 0.2 def |
8.3 ± 0.1 h |
1.6 ± 0.1 c |
4.7 ± 0.4 b |
2.7 ± 0.2 abc |
56.5 ± 2.8 bcd |
51 ± 53.8 a |
1 ± 0.4 b |
141.4 ± 57.4 cde |
|
Pressed tofu |
4.4 ± 0 i |
5.3 ± 0.2 ef |
nd |
nd |
nd |
nd |
3.9 ± 0.1 g |
nd |
0.8 ± 0 b |
14.3 ± 0.2 i |
|
Filled tofu |
15.7 ± 0.3 ab |
6.8 ± 0.1 bcd |
nd |
nd |
nd |
nd |
60.6 ± 0.2 abcd |
17 ± 0.8 b |
0.8 ± 0 b |
100.9 ± 0 ef |
|
Natto D2 |
15.6 ± 0.4 a |
9.2 ± 0.2 a |
31.5 ± 1.4 fe |
52 ± 5.1 a |
2.2 ± 0.3 cdef |
2.4 ± 0.1 abcde |
40.5 ± 1.5 e |
10.9 ± 0.9 b |
2.1 ± 0.1 ab |
166.3 ± 8.2 bc |
|
Natto D4 |
13.6 ± 0.6 ab |
7.7 ± 1.5 b |
41.2 ± 0.2 c |
41.3 ± 5.5 b |
1.8 ± 0 ef |
2.8 ± 0.3 ab |
39.4 ± 2.9 e |
11.6 ± 1.1 b |
1.9 ± 0.1 ab |
161.3 ± 8.9 bc |
|
Natto D6 |
14.6 ± 3.2 ab |
7.5 ± 1.3 bc |
58.3 ± 9 b |
43.1 ± 0.6 b |
2.2 ± 0 cdef |
1.9 ± 0.2 cde |
40.3 ± 1.5 e |
15 ± 0.4 b |
1.6 ± 0 b |
184.5 ± 2.7 ab |
|
Tempeh D2 |
10.4 ± 0.9 cdef |
6.6 ± 0.2 bcd |
8.7 ± 0.3 h |
0.1 ± 0 c |
8.7 ± 1.8 a |
3 ± 0.3 a |
5.4 ± 1 g |
11.1 ± 0.4 b |
1.7 ± 0.5 b |
55.8 ± 0.6 h |
|
Tempeh D4 |
12.4 ± 0.4 bcd |
6.2 ± 0.2 ed |
25.3 ± 1.1 fg |
0.9 ± 0 c |
3.3 ± 0.4 cd |
1.8 ± 0.1 cde |
21.5 ± 3.2 f |
7.1 ± 0 b |
1.2 ± 0.2 b |
79.7 ± 3.4 gh |
|
Tempeh D6 |
10.7 ± 0.1 cde |
4.3 ± 0.2 f |
23.9 ± 0.2 g |
1.0 ± 0.2 c |
2.9 ± 0.1 cde |
2 ± 0.1 bcde |
21 ± 3.1 f |
7.1 ± 0 b |
1.1 ± 0.1 b |
74.1 ± 3.3 gh |
|
Yogurt D0 |
7.9 ± 1.8 fgh |
5.1 ± 0.9 ef |
39.4 ± 4.2 c |
1.8 ± 0.2 c |
2.2 ± 0.2 cdef |
1.5 ± 0.2 e |
62.9 ± 2.9 abc |
14.4 ± 1.5 b |
1.9 ± 0.2 ab |
137.1 ± 1.1 cde |
|
Yogurt D2 |
6.7 ± 0.1 ghi |
5.8 ± 0 de |
37.9 ± 2.4 cde |
1.9 ± 0.2 c |
1.4 ± 1.1 f |
2.1 ± 1.1 bcde |
64.9 ± 2.6 ab |
15.6 ± 1.2 b |
2.1 ± 0.5 ab |
138.3 ± 7 cde |
|
Yogurt D4 |
10.3 ± 1.3 cdef |
5.7 ± 0.2 de |
43 ± 3.8 c |
1.8 ± 0 c |
2.3 ± 0.1 cdef |
1.8 ± 0.1 cde |
64.6 ± 1 ab |
16.1 ± 0.8 b |
2.2 ± 0.2 ab |
147.8 ± 2.7 cd |
|
Yogurt D6 |
8.9 ± 0.6 efg |
5.8 ± 0.4 de |
37.7 ± 1.5 cde |
2 ± 0.2 c |
2.1 ± 0.1 def |
1.9 ± 0.1 cde |
61.5 ± 5.2 abc |
16.5 ± 2.2 b |
2.3 ± 0.3 ab |
138.6 ± 1 cde |
|
Yogurt D8 |
9.2 ± 0.6 efg |
5.7 ± 0.2 de |
41.4 ± 4.3 c |
1.8 ± 0.1 c |
2 ± 0.1 def |
2.5 ± 0.9 abc |
60.8 ± 5.1 abcd |
15.8 ± 1 b |
2.5 ± 0.6 ab |
141.8 ± 0.5 cde |
a Data are expressed as means of triplicate experiments on dry weight basis. Values marked by the same letter within each column are not significantly different (p > 0.05). Phenolic acids: GA, gallic acid; PA, protocatechuic acid; TBA, 2,3,4-trihydroxybenzoic acid; PCD, protocatechualdehyde; HBA, p-hydroxybenzoic acid; GNA, gentistic acid; SA, syringic acid; VA, vanillic acid; VN, vanillin; CLA, chlorogenic acid; PCA + SD, p-coumaric acid + syringaldehyde; MCA + FA , m-coumaric acid + ferullic acid; SPA, sinapic acid; OCA, o-coumaric acid. ND, not detectable; RTSmilk, raw traditional soymilk; CTSmilk, cooked traditional soymilk; RSmilk slurry, raw soymilk slurry; CSmilkOF, cooked soymilk with okra and filtered.
Table 3a. Isoflavone (µg/g) profiles in fermented and non-fermented soybean products a
|
Samples |
β-glucosides |
malonylglucosides |
Acetlyglucosides |
||||||
|
|
Din |
Gin |
Gly |
MDin |
MGin |
MGly |
ADin |
AGin |
AGly |
|
Soybean |
282.6 ± 4.4 h |
536.2 ± 27.9 g |
79.9 ± 6.8 gh |
926.5 ± 5.2 a |
1813.7 ± 1.6 a |
103.4 ± 2 bc |
166.7 ± 1.2 a |
24.3 ± 0.9 fg |
34.7 ± 0.1 e |
|
RTSmilk |
205.4 ± 2.3 i |
437.4 ± 15.4 h |
69.2 ± 1.0 h |
587.1 ± 10.2 d |
1268.5 ± 15.3 e |
110.8 ± 0.8 ab |
81.1 ± 10.9 fg |
5.3 ± 0.5 k |
13.7 ± 0.7 hi |
|
CTSmilk |
402.2 ± 2.2 ef |
753.6 ± 5.9 e |
100.9 ± 1.2 ef |
394.6 ± 3 f |
813.8 ± 2.2 i |
68.6 ± 1.1 e |
75.3 ± 2.0 g |
17.6 ± 0.9 h |
60.6 ± 0.1 c |
|
RSmilk slurry |
83.3 ± 3.2 j |
233.8 ± 15.4 i |
48.6 ± 0.3 i |
796.1 ± 2.0 b |
1541 ± 2.9 b |
118.4 ± 1.7 a |
129.5 ± 12.3 c |
9.4 ± 0.6 j |
12.2 ± 0.5 hij |
|
CSmilkOF |
445.1 ± 4.0 de |
754.8 ± 4.0 e |
104.8 ± 1.7 e |
458.1 ± 3.4 ef |
937.1 ± 4.3 h |
79.3 ± 0.7 ed |
83.4 ± 1.3 fg |
26.3 ± 1.1 f |
74.5 ± 2.6 b |
|
Pressed tofu |
211.6 ± 6.3 i |
457.2 ± 19.3 h |
49.9 ± 1.3 i |
511 ± 6.2 de |
1105.2 ± 5.3 f |
73.1 ± 0.3 e |
89.3 ± 8.5 fe |
21.3 ± 0.5 g |
56.1 ± 0.9 dc |
|
Filled tofu |
349.4 ± 9.2 g |
655.9 ± 13.8 f |
88.4 ± 0.7 fg |
692.5 ± 9.2 c |
1305.6 ± 6.0 d |
91.6 ± 16.4 cd |
107.2 ± 6.4 d |
17.5 ± 4.7 h |
78.1 ± 0.8 b |
|
Natto D2 |
462.2 ± 90.7 d |
1013.6 ± 38.1 d |
130.4 ± 28.7 d |
289.2 ± 60 g |
674.9 ± 38.2 j |
65.2 ± 4.3 e |
30 ± 4.1 h |
76.4 ± 4.1 a |
53.2 ± 15.8 d |
|
Natto D4 |
392.2 ± 42.6 fg |
619.8 ± 54.2 f |
99.3 ± 12.7 ef |
700.1 ± 60.5 c |
1365.3 ± 70.1 c |
72.9 ± 18.8 e |
151.7 ± 22.3 b |
43.2 ± 2.2 d |
87.9 ± 6.5 a |
|
Natto D6 |
252.4 ± 2.3 hi |
511.4 ± 4.7 g |
67.9 ± 0.6 h |
525.7 ± 4.9 de |
1078.6 ± 10.4 g |
50.1 ± 0.5 f |
97.1 ± 0.9 de |
36.3 ± 0.3 e |
75.2 ± 0.7 b |
|
Tempeh D2 |
12.2 ± 0.8 k |
49.5 ± 2.4 j |
23.6 ± 2.1 j |
72.3 ± 3.3 h |
230.4 ± 6.9 l |
12.8 ± 3.3 ghi |
19.6 ± 0.7 hi |
50.5 ± 0.2 c |
23.2 ± 0.2 fg |
|
Tempeh D4 |
8.9 ± 1.0 k |
33.3 ± 3.0 j |
25.5 ± 4.5 j |
77.3 ± 1.5 h |
252.4 ± 2.4 k |
14.4 ± 6.2 ghi |
17.4 ± 5.5 hi |
60.4 ± 0.3 b |
26.2 ± 0.1 f |
|
Tempeh D6 |
9.5 ± 1.3 k |
35.6 ± 5.2 j |
25.7 ± 3.2 j |
79 ± 1.7 h |
256.5 ± 7.9 k |
15.3 ± 6.4 ghi |
18.6 ± 5.7 hi |
63.6 ± 0.4 b |
26.7 ± 0.2 f |
|
Yogurt D0 |
1139.5 ± 52.7 a |
1954 ± 35.7 b |
214.7 ± 25.6 ab |
114.1 ± 2.9 h |
180.4 ± 14.7 m |
10.7 ± 4.5 hi |
18.7 ± 4.5 hi |
14.4 ± 0.2 hi |
5.7 ± 2.8 j |
|
Yogurt D2 |
1126.8 ± 45.1 ab |
2003.8 ± 11.7 a |
219.2 ± 12.2 a |
79 ± 13.6 h |
131.1 ± 5.2 o |
8.1 ± 6.1 i |
14.3 ± 2.8 i |
13.4 ± 3.7 i |
8.9 ± 2.4 ij |
|
Yogurt D4 |
1031.3 ± 27.6 c |
1808.6 ± 50.9 c |
196.9 ± 4.9 c |
84.1 ± 6.5 h |
150.7 ± 2.1 n |
12.2 ± 14.5 ghi |
13.4 ± 5.2 i |
12.6 ± 1.4 ij |
10.5 ± 0.7 ij |
|
Yogurt D6 |
1101.8 ± 37.7 ab |
2003.1 ± 15.4 a |
201.1 ± 6.9 cb |
77 ± 6.5 h |
150.8 ± 1.5 n |
26.9 ± 1.4 g |
16.3 ± 3.6 hi |
13.4 ± 1.5 i |
5.8 ± 0.1 j |
|
Yogurt D8 |
1084.8 ± 25.4 b |
1957.3 ± 19.2 b |
203.2 ± 6.2 cb |
86.9 ± 8.9 h |
166.9 ± 1.3 mn |
24.8 ± 6.3 gh |
16.3 ± 4.2 hi |
13.3 ± 1.7 i |
17.5 ± 4.7 gh |
Table 3b. Isoflavone (µg/g) profiles in fermented and non-fermented soybean products a (continued)
|
Samples |
aglycones |
total individuals b |
Total isoflavones c |
||||
|
|
Dein |
Gein |
Glein |
T-Dein |
T-Gein |
T-Glein |
|
|
Soybean |
32.7 ± 2.9 l |
13.5 ± 1.6 i |
20.8 ± 1.1 e |
766.6 ± 2.1 cd |
1272.4 ± 18.7 b |
141.2 ± 3.4 cde |
2180.2 ± 19.4 ab |
|
RTSmilk |
412.0 ± 1.1 cd |
259.3 ± 8.9 d |
49.4 ± 1.2 cb |
879.6 ± 3.5 a |
1173.4 ± 14.6 ef |
155 ± 0.9 bcd |
2208.4 ± 15.1 a |
|
CTSmilk |
348.3 ± 2.3 e |
200.5 ± 6.8 e |
52.6 ± 0.3 ab |
835.3 ± 1.3 ab |
1082.5 ± 13.4 g |
182.6 ± 1.4 a |
2100.4 ± 23.6 bcd |
|
RSmilk slurry |
198.1 ± 0.8 g |
251.6 ± 5.7 d |
27.9 ± 1.7 d |
723.7 ± 4.6 d |
1179.4 ± 8.3 e |
124.3 ± 0.9 ef |
2027.3 ± 11.8 d |
|
CSmilkOF |
286.1 ± 4.9 f |
235.4 ± 2.8 d |
47.4 ± 0.1 c |
836.1 ± 6.1 ab |
1185.4 ± 10.4 de |
192.9 ± 2.2 a |
2214.4 ± 16.6 a |
|
Pressed tofu |
108.1 ± 2.6 j |
167.5 ± 3.6 f |
15.2 ± 0.5 f |
545.3 ± 13.3 ef |
1017.9 ± 18 h |
113.8 ± 1.6 f |
1677.1 ± 32.7 g |
|
Filled tofu |
127.2 ± 1.1 i |
177.1 ± 1.6 h |
18.3 ± 0.7 ef |
750.6 ± 6.7 cd |
1248.2 ± 12.8 bc |
162 ± 8.6 b |
2160.8 ± 2.1 abc |
|
Natto D2 |
149.8 ± 2.8 h |
130.4 ± 12.8 g |
16.0 ± 1.4 f |
595.2 ± 88.1 e |
1133.6 ± 11.2 f |
158.2 ± 39.8 bc |
1887.0 ± 139.1 e |
|
Natto D4 |
37.3 ± 0.5 k |
58.5 ± 4.7 h |
-0.1 ± 0.0 h |
715.4 ± 78.5 d |
1152 ± 31.1 ef |
146.3 ± 14.4 bcd |
2013.7 ± 115.2 d |
|
Natto D6 |
39 ± 0.4 k |
52.1 ± 0.5 h |
-0.1 ± 0 h |
513.2 ± 4.8 f |
930.6 ± 8.6 i |
108.5 ± 1.9 f |
1552.2 ± 14.4 h |
|
Tempeh D2 |
430.1 ± 2.5 bc |
980.8 ± 6.1 c |
53.5 ± 0.1 ab |
485.1 ± 1.1 f |
1155.7 ± 3.3 ef |
87.2 ± 1.4 g |
1728.3 ± 4.9 g |
|
Tempeh D4 |
434.7 ± 3.6 b |
1009 ± 2.7 b |
54.6 ± 2.1 a |
488.7 ± 3.7 f |
1190.6 ± 2.9 de |
92.8± 5.2 g |
1771.2 ± 0.9 fg |
|
Tempeh D6 |
451.2 ± 2.8 a |
1049.3 ± 2.4 a |
53.5 ± 4 ab |
507 ± 4.1 f |
1236.2 ± 3.4 bc |
91.6 ± 7.3 g |
1834.8 ± 3.1 ef |
|
Yogurt D0 |
27.9 ± 0.1 m |
32.4 ± 1.3 hi |
5.3 ± 0 g |
792.3 ± 31.2 bc |
1325.6 ± 27.8 a |
145.6 ± 11.7 bcd |
2263.0 ± 70.8 a |
|
Yogurt D2 |
28.4 ± 0.4 m |
32.4 ± 8.8 hi |
4.7 ± 1.3 g |
764.7 ± 26.7 cd |
1330.5 ± 9.9 a |
147.6 ± 5.4 bcd |
2242.9 ± 38.8 a |
|
Yogurt D4 |
26.3 ± 0.6 m |
35.3 ± 0.7 hi |
4.5 ± 0.2 g |
706.4 ± 17.2 d |
1223.6 ± 32 cd |
136.7 ± 10.2 ed |
2066.7 ± 59.3 bdc |
|
Yogurt D6 |
29.2 ± 0.9 m |
41.2 ± 1.9 hi |
9.2 ± 3.2 g |
750.4 ± 22.8 cd |
1348.8 ± 2.6 a |
148.9 ± 6.1 bcd |
2248.1 ± 28.4 a |
|
Yogurt D8 |
27.1 ± 0.1 m |
36.6 ± 1.2 hi |
6.2 ± 1.9 g |
742.9 ± 13.1 cd |
1324.2 ± 6.4 a |
152.6 ± 8.1 bcd |
2219.7 ± 17.1 a |
a Data are expressed as means of triplicate experiments on dry weight basis. Values marked by the same letter within each soybean in each column are not significantly different (p > 0.05). Din, daidzin; Gin, genistin; Gly, glycitin; MDin, malonyldaidzin; MGin, malonylgenistin; MGly, malonylglycitin; ADin, acetyldaidzin; AGin, acetylgenistin; AGly, acetylglycitin; Dein, daidzein; Gein, genistein; Glein, glycitein; T-Dein, subtotal daidzein; T-Gein, subtotal genistein; T-Glein, subtotal glycitein. ND, not detected; RTSmilk, raw traditional soymilk; CTSmilk, cooked traditional soymilk; RSmilk slurry, raw soymilk slurry; CSmilkOF, cooked soymilk with okra and filtered. b Subtotal individuals = moles of isoflavone × molecular weight of aglycone form isoflavone. c Total isoflavones = sum of subtotal individuals of aglycones.
Table 4. Correlations between antioxidant activities and phenolic compounds in fermented and non-fermented soybean products a
|
|
Fermented products (N=33) a |
Non-fermented products (N=21) a |
|||||
|
Phenolic compounds |
DPPH |
FRAP |
ORAC |
Phenolic compounds |
DPPH |
FRAP |
ORAC |
|
TPC |
-0.52 |
-0.34 |
0.85 b |
TPC |
0.68 c |
0.26 |
0.83 c |
|
TFC |
0.66 c |
0.29 |
0.01 |
CTC |
0.71 |
-0.31 |
0.90 c |
|
CTC |
-0.19 |
-0.36 |
0.80 b |
F-PA |
0.76 c |
-0.04 |
0.61 |
|
F-PA |
-0.01 |
0.09 |
0.88 b |
F-TBA |
0.77 c |
-0.13 |
0.64 |
|
F-PCD |
-0.51 |
-0.36 |
0.88 b |
F-GNA |
0.42 |
-0.02 |
0.78 c |
|
F-HBA |
-0.54 |
-0.4 |
0.82 b |
F-SA |
0.7 |
0.05 |
0.80 c |
|
F-GNA |
-0.67 c |
-0.27 |
0.16 |
F-VN |
0.68 |
-0.21 |
0.82 c |
|
F-SA |
-0.41 |
-0.11 |
0.93 b |
T-GNA |
0.78 c |
-0.06 |
0.6 |
|
F-Ben |
-0.21 |
0.01 |
0.96 b |
T-VA |
0.76 c |
-0.16 |
0.80 c |
|
F-CLA |
-0.68 c |
-0.34 |
0.58 |
T-Ben |
0.62 |
0.2 |
0.36 |
|
F-TP |
-0.5 |
-0.02 |
0.69 c |
T-CLA |
0.94 c |
0.38 |
0.64 |
|
T-GA |
-0.12 |
-0.39 |
0.75 c |
Gly |
0.77 c |
-0.06 |
0.56 |
|
T-PA |
-0.18 |
-0.17 |
0.76 c |
MGly |
-0.01 |
0.79 c |
0.02 |
|
T-TBA |
-0.61 c |
-0.80 c |
0 |
Dein |
0.27 |
-0.14 |
0.78 c |
|
T-PCD |
-0.42 |
-0.56 |
0.75 c |
Glein |
0.54 |
-0.18 |
0.85 c |
|
T-HBA |
0.89 b |
0.64 c |
-0.1 |
T-Dein |
0.73 |
0.37 |
0.79 c |
|
T- Ben |
-0.70 c |
-0.66 c |
0.33 |
T-Gein |
0.5 |
0.83 c |
0.16 |
|
T-CFA |
-0.18 |
-0.74 c |
0.23 |
T-Glein |
0.78 c |
-0.06 |
0.77 c |
|
T-CLA |
0.28 |
0.1 |
-0.26 |
TI |
0.78 c |
0.59 |
0.65 |
|
T-MCA |
-0.64 c |
-0.67 c |
0.1 |
|
|
|
|
|
TP |
-0.64 c |
-0.46 |
0.03 |
|
|
|
|
|
Gin |
-0.54 |
0.04 |
0.84 b |
|
|
|
|
|
MDin |
-0.23 |
-0.69 c |
0.32 |
|
|
|
|
|
MGin |
-0.2 |
-0.66 c |
0.39 |
|
|
|
|
|
MGly |
-0.39 |
-0.52 |
0.63 c |
|
|
|
|
|
ADin |
-0.11 |
-0.65 c |
0.12 |
|
|
|
|
|
AGin |
0.09 |
0.06 |
0.68 c |
|
|
|
|
|
AGly |
-0.14 |
-0.61 c |
0.47 |
|
|
|
|
|
T-Glein |
-0.64 c |
-0.2 |
0.18 |
|
|
|
|
a Values were from triplicate determinations on fermented (natto day 2, 4, 6; tempeh day 2, 4, 6; yogurt day 0, 2, 4, 6, 8) and non-fermented (soybean; traditional raw soymilk; traditional cooked soymilk; raw soymilk slurry; cooked soymilk with okra and filtered; pressed tofu; filled tofu) soybean products . b The correlation is significant at p = 0.0001 (two-tailed). c The correlation is significant at p = 0.05 (two-tailed). TPC, total phenolic content; CTC, condensed tannin content; F, free phenolic acid analysis (F-GA, free gallic acid, F-PA, free protocatechuic acid; F- PCD, protocatechualdehyde; F-TBA, free 2,3,4-trihydroxybenzoic acid; F-GNA, free gentistic acid; F-HBA, free p-hydroxybenzoic acid; F-SA, free syringic acid; F-VA, free vanillic acid; F-Ben, Free subtotal benzoics; F-CLA, free chlorogenic acid; F-PCA+SD, free p-coumaric acid + syringaldehyde F-Cin, Free subtotal cinnamics; F-TP, free total phenolics content) ; Total, conjugated phenolic acid analysis (T-GA, total gallic acid, T-HBA, total p-hydroxybenzoic acid; T-GNA, gentistic acid; T-SA, total syringic acid; T-VA, conjugated vanillic acid; T-Ben, subtotal benzoics; T-CLA, total chlorogenic acid; T-PCA+SD, total p-coumaric acid + syringaldehyde T-Cin, subtotal cinnamics; TP, total phenolics content, sum of all individual acids); Gin, genistin, Gly, glycitin, MDin, malonyldaidzin; MGin, malonylgenistin; MGly, malonylglycitin; ADin, acetyldaidzin; AGin, acetylgenistin; AGly, acetylglycitin; Dein, daidzein; Gein, genistein; T-Dein, subtotal daidzein; T-Gein, subtotal genistein; T-Glein, subtotal glycitein; TI, total isoflavones.
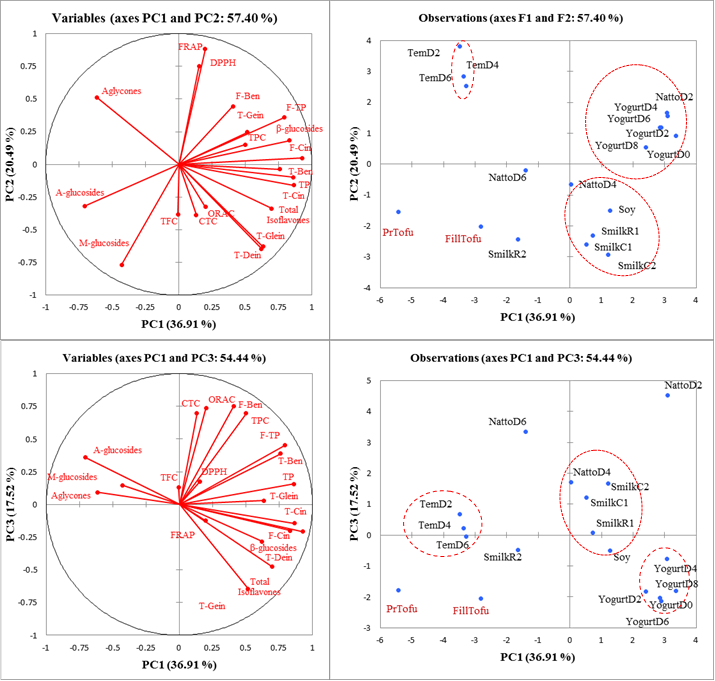
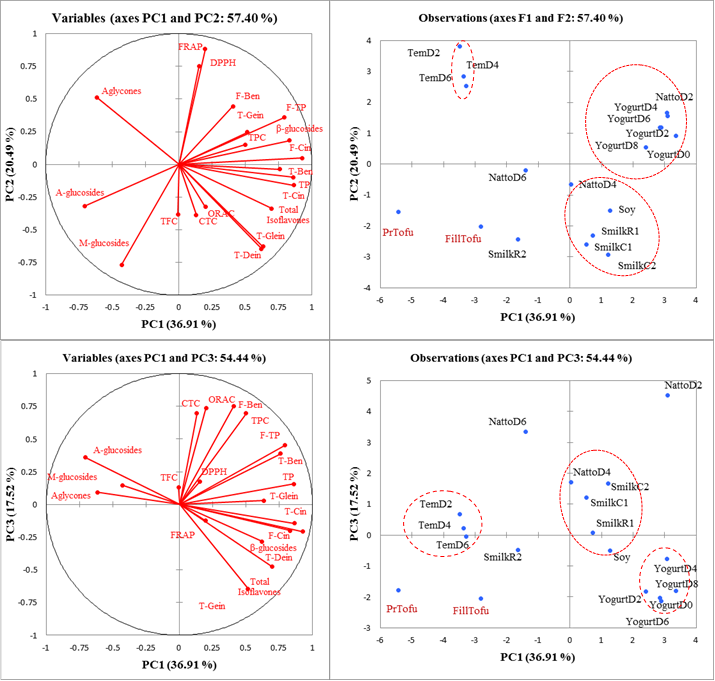
1. Tsukamoto Y, Ichise H, Kakuda H, Yamaguchi M. Intake of fermented soybean ( natto ) increases circulating vitamin K 2. (menaquinone-7) and γ-carboxylated osteocalcin concentration in normal individuals. Journal of Bone and Mineral Metabolism. 2000;18(4):216-22.
2. Liu J, Chang SKC, Wiesenborn D. Antioxidant Properties of Soybean Isoflavone Extract and Tofu in Vitro and in Vivo. Journal of Agricultural and Food Chemistry. 2005;53(6):2333-40.
3. Park D, Huang T, Frishman WH. Phytoestrogens as Cardioprotective Agents. Cardiology in Review. 2005;13(1):13-7.
4. Yao F-R, Sun C-W, Chang SKC. Morton Lentil Extract Attenuated Angiotensin II-Induced Cardiomyocyte Hypertrophy via Inhibition of Intracellular Reactive Oxygen Species Levels in Vitro. Journal of Agricultural and Food Chemistry. 2010;58(19):10382-8.
5. Valko M, Leibfritz D, Moncol J, Cronin MTD, Mazur M, Telser J. Free radicals and antioxidants in normal physiological functions and human disease. The International Journal of Biochemistry & Cell Biology. 2007;39(1):44-84.
6. Pratt DE. Natural Antioxidants of Soybeans and Other Oil-Seeds. Autoxidation in Food and Biological Systems: Springer US; 1980. p. 283-93.
7. Jackson CJC, Dini JP, Lavandier C, Rupasinghe HPV, Faulkner H, Poysa V, et al. Effects of processing on the content and composition of isoflavones during manufacturing of soy beverage and tofu. Process Biochemistry. 2002;37(10):1117-23.
8. Tan Y, Chang SKC, Zhang Y. Innovative Soaking and Grinding Methods and Cooking Affect the Retention of Isoflavones, Antioxidant and Antiproliferative Properties in Soymilk Prepared from Black Soybean. Journal of Food Science. 2016;81(4):H1016-H23.
9. Xu B, Chang SKC. Isoflavones, Flavan-3-ols, Phenolic Acids, Total Phenolic Profiles, and Antioxidant Capacities of Soy Milk As Affected by Ultrahigh-Temperature and Traditional Processing Methods. Journal of Agricultural and Food Chemistry. 2009;57(11):4706-17.
10. Zhang Y, Chang SKC, Liu Z. Isoflavone Profile in Soymilk as Affected by Soybean Variety, Grinding, and Heat-Processing Methods. Journal of Food Science. 2015;80(5):C983-C8.
11. Kim EH, Kim SH, Chung JI, Chi HY, Kim JA, Chung IM. Analysis of phenolic compounds and isoflavones in soybean seeds (Glycine max (L.) Merill) and sprouts grown under different conditions. European Food Research and Technology. 2005;222(1-2):201-8.
12. Kumari S, Chang SKC. Effect of Cooking on Isoflavones, Phenolic Acids, and Antioxidant Activity in Sprouts of Prosoy Soybean (Glycine max). Journal of Food Science. 2016;81(7):C1679-C91.
13. Xu B, Chang SKC. Reduction of antiproliferative capacities, cell-based antioxidant capacities and phytochemical contents of common beans and soybeans upon thermal processing. Food Chemistry. 2011;129(3):974-81.
14. Xu B, Chang SKC. Effect of soaking, boiling, and steaming on total phenolic contentand antioxidant activities of cool season food legumes. Food Chemistry. 2008;110(1):1-13.
15. Chung I-M, Seo S-H, Ahn J-K, Kim S-H. Effect of processing, fermentation, and aging treatment to content and profile of phenolic compounds in soybean seed, soy curd and soy paste. Food Chemistry. 2011;127(3):960-7.
16. Hur SJ, Lee SY, Kim Y-C, Choi I, Kim G-B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chemistry. 2014;160:346-56.
17. Huynh NT, Van Camp J, Smagghe G, Raes K. Improved release and metabolism of flavonoids by steered fermentation processes: a review. Int J Mol Sci. 2014;15(11):19369-88.
18. McCue P, Horii A, Shetty K. Solid-state bioconversion of phenolic antioxidants from defatted soybean powders by Rhizopus oligosporus: role of carbohydrate-cleaving enzymes. Journal of Food Biochemistry. 2003;27(6):501-14.
19. Martins S, Mussatto SI, Martínez-Avila G, Montañez-Saenz J, Aguilar CN, Teixeira JA. Bioactive phenolic compounds: Production and extraction by solid-state fermentation. A review. Biotechnology Advances. 2011;29(3):365-73.
20. Haron H, Raob N. Changes in macronutrient, total phenolic and anti-nutrient contents during preparation of tempeh. Nutrition & Food Sciences. 2014.
21. Hati S, Vij S, Singh BP, Mandal S. β -Glucosidase activity and bioconversion of isoflavones during fermentation of soymilk. Journal of the Science of Food and Agriculture. 2014;95(1):216-20.
22. Marazza JA, LeBlanc JG, de Giori GS, Garro MS. Soymilk fermented with Lactobacillus rhamnosus CRL981 ameliorates hyperglycemia, lipid profiles and increases antioxidant enzyme activities in diabetic mice. Journal of Functional Foods. 2013;5(4):1848-53.
23. Juan M-Y, Chou C-C. Enhancement of antioxidant activity, total phenolic and flavonoid content of black soybeans by solid state fermentation with Bacillus subtilis BCRC 14715. Food Microbiology. 2010;27(5):586-91.
24. Shin E-C, Lee JH, Hwang CE, Lee BW, Kim HT, Ko JM, et al. Enhancement of total phenolic and isoflavone-aglycone contents and antioxidant activities during Cheonggukjang fermentation of brown soybeans by the potential probiotic Bacillus subtilis CSY191. Food Science and Biotechnology. 2014;23(2):531-8.
25. Dajanta K, Apichartsrangkoon A, Chukeatirote E. Antioxidant Properties and Total Phenolics of Thua Nao (a Thai Fermented Soybean)as Affected by Bacillus-fermentation. Journal of Microbial & Biochemical Technology. 2011;03(04).
26. Toda T, Sakamoto A, Takayanagi T, Yokotsuka K. Changes in Isoflavone Compositions of Soybean Foods during Cooking Process. Food Science and Technology Research. 2000;6(4):314-9.
27. Xu B, Chang SKC. Total Phenolics, Phenolic Acids, Isoflavones, and Anthocyanins and Antioxidant Properties of Yellow and Black Soybeans As Affected by Thermal Processing. Journal of Agricultural and Food Chemistry. 2008;56(16):7165-75.
28. Handa CL, Zhang Y, Kumari S, Xu J, Ida EI, Chang SK. Comparative Study of Angiotensin I-Converting Enzyme (ACE) Inhibition of Soy Foods as Affected by Processing Methods and Protein Isolation. Processes. 2020;8(8):978.
29. Zhang Y, Guo S, Liu Z, Chang SKC. Off-Flavor Related Volatiles in Soymilk As Affected by Soybean Variety, Grinding, and Heat-Processing Methods. Journal of Agricultural and Food Chemistry. 2012;60(30):7457-62.
30. Meng S, Chang S, Gillen AM, Zhang Y. Protein and quality analyses of accessions from the USDA soybean germplasm collection for tofu production. Food Chemistry. 2016;213:31-9.
31. Donkor ON, Henriksson A, Vasiljevic T, Shah NP. Rheological Properties and Sensory Characteristics of Set-Type Soy Yogurt. Journal of Agricultural and Food Chemistry. 2007;55(24):9868-76.
32. Wang Y-C, Yu R-C, Chou C-C. Growth and survival of bifidobacteria and lactic acid bacteria during the fermentation and storage of cultured soymilk drinks. Food Microbiology. 2002;19(5):501-8.
33. Xu B, Chang SKC. Characterization of Phenolic Substances and Antioxidant Properties of Food Soybeans Grown in the North Dakota−Minnesota Region. Journal of Agricultural and Food Chemistry. 2008;56(19):9102-13.
34. Collison MW. Determination of total soy isoflavones in dietary supplements, supplement ingredients, and soy foods by high-performance liquid chromatography with ultraviolet detection: collaborative study. Journal of AOAC International. 2008;91(3):489-500.
35. Cho K, Hong S, Math R, Lee J, Kambiranda D, Kim J, et al. Biotransformation of phenolics (isoflavones, flavanols and phenolic acids) during the fermentation of cheonggukjang by Bacillus pumilus HY1. Food Chemistry. 2009;114(2):413-9.
36. Prior RL, Wu X, Schaich K. Standardized Methods for the Determination of Antioxidant Capacity and Phenolics in Foods and Dietary Supplements. Journal of Agricultural and Food Chemistry. 2005;53(10):4290-302.
37. Lee SJ, Yan W, Ahn JK, Chung IM. Effects of year, site, genotype and their interactions on various soybean isoflavones. Field Crops Research. 2003;81(2-3):181-92.
38. Iwashita K, Shimoi H, Ito K. Extracellular soluble polysaccharide (ESP) from Aspergillus kawachii improves the stability of extracellular β-gluocosidases (EX-1 and EX-2) and is involved in their localization. Journal of Bioscience and Bioengineering. 2001;91(2):134-40.
39. Pyo Y-H, Lee T-C, Lee Y-C. Enrichment of bioactive isoflavones in soymilk fermented with β-glucosidase-producing lactic acid bacteria. Food Research International. 2005;38(5):551-9.
40. Georgetti SR, Vicentini FTMC, Yokoyama CY, Borin MF, Spadaro ACC, Fonseca MJV. Enhancedin vitroandin vivoantioxidant activity and mobilization of free phenolic compounds of soybean flour fermented with different β-glucosidase-producing fungi. Journal of Applied Microbiology. 2009;106(2):459-66.
41. Torino MI, Limón RI, Martínez-Villaluenga C, Mäkinen S, Pihlanto A, Vidal-Valverde C, et al. Antioxidant and antihypertensive properties of liquid and solid state fermented lentils. Food Chemistry. 2013;136(2):1030-7.
42. Dueñas M, Fernández D, Hernández T, Estrella I, Muñoz R. Bioactive phenolic compounds of cowpeas (Vigna sinensisL). Modifications by fermentation with natural microflora and withLactobacillus plantarumATCC 14917. Journal of the Science of Food and Agriculture. 2004;85(2):297-304.
43. Ng TB, Liu F, Wang ZT. Antioxidative activity of natural products from plants. Life Sciences. 2000;66(8):709-23.
44. Jung S, Murphy PA, Sala I. Isoflavone profiles of soymilk as affected by high-pressure treatments of soymilk and soybeans. Food Chemistry. 2008;111(3):592-8.
45. Yuan S, Chang SK, Liu Z, Xu B. Elimination of trypsin inhibitor activity and beany flavor in soy milk by consecutive blanching and ultrahigh-temperature (UHT) processing. Journal of agricultural and food chemistry. 2008;56(17):7957-63.
46. Wang H-J, Murphy PA. Mass Balance Study of Isoflavones during Soybean Processing. Journal of Agricultural and Food Chemistry. 1996;44(8):2377-83.
47. Kudou S, Tsuizaki I, Uchida T, Okubo K. Purification and some properties of soybean saponin hydrolase from Aspergillus oryzae KO-2. Agricultural and Biological Chemistry. 1991;55(1):31-6.
48. Setchell KDR, Brown NM, Zimmer-Nechemias L, Brashear WT, Wolfe BE, Kirschner AS, et al. Evidence for lack of absorption of soy isoflavone glycosides in humans, supporting the crucial role of intestinal metabolism for bioavailability. The American Journal of Clinical Nutrition. 2002;76(2):447-53.
49. Izumi T, Piskula MK, Osawa S, Obata A, Tobe K, Saito M, et al. Soy Isoflavone Aglycones Are Absorbed Faster and in Higher Amounts than Their Glucosides in Humans. The Journal of Nutrition. 2000;130(7):1695-9.
50. Rostagno MA, Villares A, Guillamón E, García-Lafuente A, Martínez JA. Sample preparation for the analysis of isoflavones from soybeans and soy foods. Journal of Chromatography A. 2009;1216(1):2-29.
51. Cheng K-C, Wu J-Y, Lin J-T, Liu W-H. Enhancements of isoflavone aglycones, total phenolic content, and antioxidant activity of black soybean by solid-state fermentation with Rhizopus spp. European Food Research and Technology. 2013;236(6):1107-13.
52. Choi H-J, Cheigh C-I, Kim S-B, Lee J-C, Lee D-W, Choi S-W, et al. Weissella kimchii sp. nov., a novel lactic acid bacterium from kimchi. International Journal of Systematic and Evolutionary Microbiology. 2002;52(2):507-11.
53. Kuo L-C, Cheng W-Y, Wu R-Y, Huang C-J, Lee K-T. Hydrolysis of black soybean isoflavone glycosides by Bacillus subtilis natto. Applied Microbiology and Biotechnology. 2006;73(2):314-20.
The Roles of Choline in Maintaining Optimal Health
The Effects of Intake of Bread with Treated Corn Bran Inclusion on Postprandial Glyceamic Response
Food Waste throughout the Food Production Continuum – Water Food and Energy Nexus
Use of Lightly Potassium-Enriched Soy Sauce at Home Reduced Urinary Sodium-to-Potassium Ratio
A Different Type of Critical Migration
PRECEDE: A Conceptual Model to Assess Immigrant Health
Walking Together: Supporting Indigenous Student Success in University
Cosmetic Surgery and Body Image in Race/Ethnic Minorities

Our articles most useful
Adrian Taylor and Marica Bakovic*
Published : June 21, 2019
Journal of Food & Nutritional Sciences
Zhimin Cui, Lynne Kennedy, Weili Li*
Published : September 30, 2019
Journal of Food & Nutritional Sciences
Elena Castell-Perez*, Rosana G. Moreira, Hal S. Knowles, III
Published : October 01, 2019
Journal of Food & Nutritional Sciences
Nagako Okuda, Makoto Miura, Kazuyoshi Itai, Takuya Morikawa, Junko Sasaki, Tamami Asanuma, Mikako Fujii, Akira Okayama
Published : March 27, 2019
Journal of Food & Nutritional Sciences