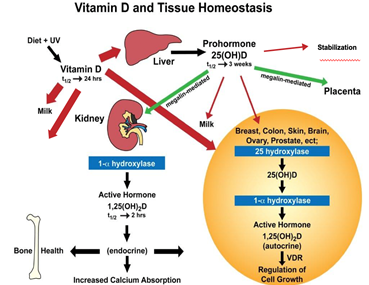
To understand vitamin D’s role during pregnancy, lactation and early infancy, we must first understand what vitamin D is, and its sources. Vitamin D is a preprohormone that is created in the epidermal layer of the skin following the exposure of 7-dehyrocholesterol to ultraviolet B light, within a specific wavelength between 290-315 nanometers [1], producing previtamin D. Through a thermal conversion in the skin, the previtamin D is converted into cholecalciferol or vitamin D3 and transported systemically via either vitamin D binding protein or albumin (see Figure 1 below). Once in the liver, vitamin D is converted through a 25-hydroxylation to 25-hydroxy-vitamin D (25(OH)D), where it then circulates throughout the body attached mainly to VDBP or albumin [2, 3]. It is converted by the proximal tubules in the kidney to the active hormone 1,25-dihydroxy-vitamin (1,25(OH)2D) with a hydroxylation at the 1-alpha position that is megalin-mediated [4]. This conversion also is regulated by parathyroid hormone. When there is vitamin D deficiency in the body, circulating PTH increases [2, 3]. Not limited to the kidneys, there are receptors to vitamin D moieties throughout the body, and include immune cells, such as dendritic cells and monocytes/macrophages and lymphocytes [6-10], where 1,25(OH)2D can be synthesized within the nuclear membrane.
*Corresponding Author:
Carol L. Wagner, MD
Shawn Jenkins Children’s Hospital, 10 McClennan Banks Drive, MSC 915, Charleston, SC 29425, USA, Tel: 843-792-2112; Fax: 843-792-8801; E-mail: wagnercl@musc.edu
To understand vitamin D’s role during pregnancy, lactation and early infancy, we must first understand what vitamin D is, and its sources. Vitamin D is a preprohormone that is created in the epidermal layer of the skin following the exposure of 7-dehyrocholesterol to ultraviolet B light, within a specific wavelength between 290-315 nanometers [1], producing previtamin D. Through a thermal conversion in the skin, the previtamin D is converted into cholecalciferol or vitamin D3 and transported systemically via either vitamin D binding protein or albumin (see Figure 1 below). Once in the liver, vitamin D is converted through a 25-hydroxylation to 25-hydroxy-vitamin D (25(OH)D), where it then circulates throughout the body attached mainly to VDBP or albumin [2, 3]. It is converted by the proximal tubules in the kidney to the active hormone 1,25-dihydroxy-vitamin (1,25(OH)2D) with a hydroxylation at the 1-alpha position that is megalin-mediated [4]. This conversion also is regulated by parathyroid hormone. When there is vitamin D deficiency in the body, circulating PTH increases [2, 3]. Not limited to the kidneys, there are receptors to vitamin D moieties throughout the body, and include immune cells, such as dendritic cells and monocytes/macrophages and lymphocytes [6-10], where 1,25(OH)2D can be synthesized within the nuclear membrane.
One of 1,25(OH)2D’s roles is to maintain calcium homeostasis. With lower 1,25(OH)2D, there is decreased absorption of calcium from the intestines, resulting in a rise in PTH, that affects osteoclasts, necessitating the demineralization of bone to maintain normal circulating calcium levels. Mellanby’s discovery in 1919 that there was something missing in the diets of children with rickets that could be cured with cod liver oil then led McCollum in 1920 to conduct experiments that showed it was not vitamin A but rather a “new” vitamin that he called “vitamin D” that was in cod liver oil, which cured rickets [11]. The discovery by Huldshinsky in 1919 that a reaction was produced in the skin following the use of mercury vapor lights to treat children with rickets, where exposure to one arm led to improvement in bony rachitic changes in the contralateral arm was a startling finding at the time, and suggested that a compound was synthesized in the light-exposed arm that could circulate and affect other sites [12, 13].
While vitamin D can be obtained from the diet, the main source in older children and adults is via sunlight exposure. The average Western diet provides about 200 IU vitamin D per day whereas thousands of IU can be generated within minutes of full sunlight exposure, during summer months or near the equator, with diminishing synthesis during late fall and winter at higher latitudes, hence making individuals living at higher latitudes at greatest risk of vitamin D deficiency. Sunscreen blocks the synthesis of vitamin D [14-17]. Those with darker pigment have an excellent filter against the ravages of ultraviolet light but require more UV-B to penetrate the melanin for conversion of 7-dehyrocholesterol to previtamin D [18, 19]. This is true across the lifespan, with the exception that young infants in most societies have less sunlight exposure due to the fragility of the skin to sun exposure [20], and therefore, are dependent on the transfer of vitamin D in the breast milk or via formula that is fortified with vitamin D [21, 22].
During pregnancy, the fetus requires transfer of 25(OH)D across the placenta to synthesize 1,25(OH)2D [21, 23]. In comparison, during lactation, it is the parent compound vitamin D3 (or less commonly the plant or fungi-derived ergocalciferol or D2) that crosses into the breast milk [21, 22, 24]. If a mother is vitamin D deficient, then during pregnancy, her fetus will be deficient [21, 23]. Similarly, during lactation, if a mother is vitamin D deficient, then her milk will be vitamin D deficient [25].
Why is vitamin D important? Clearly, earlier studies demonstrated its role in maintaining calcium homeostasis, and indirectly, bone integrity. While there were reports of individuals with vitamin D deficiency having increased respiratory infections [26-29], it was not until the 21st century that vitamin D’s role in enabling both innate [9] and adaptive immunity [30-32] was appreciated.
There is no question that vitamin D is important for early life, and by extension, for a woman during her pregnancy as the sole source of vitamin D for the developing fetus, and as the main source for her recipient breastfeeding infant [21, 33]. As mentioned earlier, there is transfer of vitamin D in the form of 25-hydroxy-vitamin D (25-OH-D) across the placenta which the fetus converts to 1,25(OH)2D in the fetal kidneys and in other tissues where 1-alpha-hydroxylase is present [34]. During lactation, it is the parent vitamin D3 (or less commonly D2) that is extruded into the breast milk and ingested by the recipient breastfeeding infant; little 25(OH)D or 1,25(OH)2D crosses into breast milk [22, 24].
There has been an emphasis through the years on the vitamin D’s endocrine role in calcium and skeletal metabolism. There has been much interest in vitamin D and certainly heated discussions about the nonskeletal role of vitamin D—that vitamin D might be the link to explaining prematurity and adverse pregnancy outcomes such as preeclampsia, but randomized controlled trials, never powered for this intention, show variable effect—in some cases a strong, positive effect [23, 35-50], and in other cases, little effect [51, 52]. Recommendation of vitamin D supplementation at whatever dose—400 international units (IU) or 5000 IU—is left to the discretion of the health care provider. There is no consensus statement by the American College of Obstetricians and Gynecologists about vitamin D supplementation during pregnancy [53, 54]. The recommendation by the Academy of Pediatrics for the breastfeeding infant is supplementation of the infant alone with no mention of supplementation of the lactating woman herself except to mention that maternal deficiency leads to lower vitamin D concentrations in the breast milk, increasing the risk of deficiency in the recipient breastfeeding infant [55]. With confusing and opposing views, what are the issues and how can one rectify this conundrum?
Our tendency to be excited about the latest new drug or therapy heralded a plethora of vitamin D supplementation studies conducted during pregnancy and lactation. Overwhelmingly, those women must at risk for vitamin D deficiency in the US during pregnancy and lactation are Black American and Hispanic women, underserved minorities, who also have higher rates of adverse pregnancy outcomes than white women [56, 57]. In other regions of the world, women living at higher latitudes or who have darker skin pigmentation and limited sunlight exposure are at greatest risk of vitamin D deficiency [58], and it is those women who show the greatest improvement in health and pregnancy outcomes when dosed with vitamin D to achieve a total circulating 25(OH)D greater than 40 ng/mL (100 nmol/L) [45], the point at which conversion of 25(OH)D to 1,25(OH)2D is optimized, the only time during the lifecycle [23].
Vitamin D has both direct and indirect effects on the immune system—promoting T regulatory cell differentiation while inhibiting differentiation of Th1 and Th17 cells, impairing developing and function of B cells, and reducing monocyte activation [59, 60]. Deficiency states affect the balance of these processes: Infants with vitamin D deficiency associate strongly with respiratory infections, chronic lung diseases such as bronchopulmonary dysplasia (BPD) and have a higher risk of respiratory syncytial virus (RSV) infection during the 1st year of life if they are found to be vitamin D deficient at birth [26, 27, 61-63]. More recently, a lower risk of asthma at 3 years of age in offspring [64-66] was demonstrated with improved maternal vitD status during pregnancy [67, 68]. In another study of 28 infants who were examined <4 d after their first allergic reaction and age-matched controls all <9 months, infants who developed cow milk protein allergy (CMPA) had decreased T regulatory (Treg) cell counts (which correlated with decreased serum 25(OH)D) and increased frequency of IL4-secreting CD4 T cells compared to controls. Values of Tregs, IL4-secreting cells and vitD are good predictors of CMPA diagnosis. Basal 25(OH)D in CMPA infants also predicted those CMPA patients developing spontaneous tolerance in the first year [69]. Thus, emerging evidence points to serious consequences of both acute and chronic vitamin D deprivation, including the implications of vitamin D deficiency during early infant development [26, 28, 29, 36-38, 69-95]. Observational trials show adverse health effects associated with vitamin D deficiency, but clinical trial results are less clear given the variability in dosing, maternal baseline vitamin D status, and maternal BMI affecting outcomes.
The effect of maternal supplementation during lactation has been less well studied. In two pilot studies along with a larger NICHD trial that our group led, we sought to determine the optimal vitamin D supplementation dose for lactating women. We proposed that if a mother was vitamin D replete, her infant would not require vitamin D supplementation. In our randomized clinical trial, 216 mother-infant exclusively breastfeeding pairs were randomized to mother and infant 400 IU/day or mother alone 6400 IU/day and infant 0 IU/day. 148 completed the study to visit #4 and 95 to visit #7 [25, 96]. Compared to 400 IU vitD/day, 6400 IU/day safely and significantly increased maternal vitD and 25-hydroxyvitamin D (25(OH)D; expressed in nmol/L) from baseline (p<0.0001), independent of race. 25(OH)D concentration did not differ in breastfeeding infants in the 400 IU group directly receiving supplement compared to infants in the 6400 IU-maternal supplemented only group. Across visits there were no differences in maternal or infant serum calcium, creatinine, phosphorus, or in urinary calcium/creatinine ratios as a function of treatment group. Safety was further established by comparing maternal safety parameters in postpartum lactating and nonlactating women and their infants [97].
Data from these trials suggested that maternal vitamin D supplementation with 6400 IU/day safely repletes breast milk with adequate vitamin D to satisfy the requirement of the breastfeeding infant and offers an alternate strategy to direct infant vitamin D supplementation. These were important findings given that a major limiting factor of supplementing vitamin D directly to the infant is a higher rate of noncompliance, even more severe among minority and poor communities [98-101]. Other studies have replicated our findings using similar [102] as well as bolus dosing [103, 104], particularly applied in populations where there may be compliance issues. We also have shown that vitamin D status correlates with VDBP genotype with a greater response to supplementation in those with the 1s allele as opposed to the 1f allele [105]. Both VDR and VDBP genotype may play a role in infant response to exogenous vitamin D via mother’s milk and/or oral supplement, and this remains to be studied. Much remains to be done to determine if there are differences in the effect of maternal vitamin D status on the immune signature of her milk, and likewise, on the recipient infant’s immune status.
Along with clinical trials, there emerged during the last two decades an exponential number of basic science and translational research endeavors that have demonstrated vitamin D’s role beyond calcium—that of the immune system, affecting both innate and adaptive immunity [3, 32, 106]. There are direct effects of vitamin D that are more easily ascertained in the laboratory where various factors are controlled then in the clinical arena. Laboratory studies elucidate the role that vitamin D plays in maintaining health. In target cells, tightly regulated 1,25(OH)2D activates the vitamin D receptor (VDR), a nuclear transcription factor, that then binds complexed with RXR to a vitamin D response element on DNA, a binding site located ~3000 times throughout the human genome. The activated VDR directly regulates expression of hundreds of known genes involved in diverse physiological processes, including calcium homeostasis, immune regulation, growth/development, redox balance, metabolism, epigenetic control, cell signaling and proliferation [47, 107-111].
We and others have shown that maternal vitamin D status during pregnancy and lactation affects DNA methylation and plays a critical role in metabolic processes and immune development and function [112-114] as well as epigenetic clock differences that might affect aging and maturation [47]. We tested new epigenetic clocks developed for neonates among a multiethnic population, and tested the hypothesis that maternal vitamin D supplementation would slow down the epigenetic gestational age acceleration (GAA) in newborn babies [47]. Maternal vitamin D3 supplementation appears to slow down the epigenetic gestational aging process in African American neonates.
Additional support of the effect of maternal vitamin D status on both maternal and infant epigenome comes from Anderson, et al., [113] who showed differences in both maternal and infant DNA methylation patterns by treatment group (3800 IU/day vs placebo). Associated gene clusters showed strongest biologic relevance for cell migration/motility and cellular membrane function at birth and cadherin signaling and immune function at postpartum. Breastfed 4–6-week-old infants of intervention mothers showed DNA methylation gain and loss in 217 and 213 CpGs, respectively, compared to controls. Genes showing differential methylation mapped most strongly to collagen metabolic processes and regulation of apoptosis. In another study by Chen et al involving obese African American participants, 4000 IU vitD3/day was associated with a decrease by 1.85 years compared to placebo using the Horrath age calculator [115]. Such findings support the premise that vitamin D has profound effects on genetic and epigenetic determinants and support the role that vitamin D status has on clinical health.
Part of the problem in demonstrating vitamin D’s clinical effect, which has been discussed in the literature [5, 116], is that the application of principles of pharmaceutical clinical trials to nutrient clinical trials is faulty [116]—in the former, a drug trial, the recipient of said drug starts with a preexisting blood concentration of zero; with nutrient and endocrine studies, there is some definable concentration in the blood of that nutrient or hormonal precursor—typically. The second issue is that blood levels do not automatically translate into what is seen at the tissue level [5]. Dosing and the range given to a particular participant in a clinical trial will be affected by specific binding proteins genotype—in the case of vitamin D—its vitamin D binding protein allele that impacts how tightly the vitamin D moiety is bound to the protein [105]. Other factors such as body mass index and the starting point of vitamin D status also affect circulating levels of vitamin D and its metabolites [52].
As we attempt to understand vitamin D’s role in the body, particularly during times of extreme growth such as pregnancy and lactation and early childhood, it is important to consider those factors beyond the science that may be affecting our interpretation of clinical data. We must seek to understand what is lacking in our clinical trial that does not explain what we see in the laboratory or in observational studies. The context in which we live and our desire for a simple answer drives us, yet a simple answer about vitamin D is elusive. Beyond the sound and fury over vitamin D clinical trials, one must focus on the physiological aspects of vitamin D to offer a glimpse at how vitamin D contributes to homeostasis and well-being in both the mother and her developing fetus during pregnancy that continues during lactation and early infancy.
REFERENCES
The Roles of Choline in Maintaining Optimal Health
The Effects of Intake of Bread with Treated Corn Bran Inclusion on Postprandial Glyceamic Response
Food Waste throughout the Food Production Continuum – Water Food and Energy Nexus
Use of Lightly Potassium-Enriched Soy Sauce at Home Reduced Urinary Sodium-to-Potassium Ratio
A Different Type of Critical Migration
PRECEDE: A Conceptual Model to Assess Immigrant Health
Walking Together: Supporting Indigenous Student Success in University
Cosmetic Surgery and Body Image in Race/Ethnic Minorities

Our articles most useful
Adrian Taylor and Marica Bakovic*
Published : June 21, 2019
Journal of Food & Nutritional Sciences
Zhimin Cui, Lynne Kennedy, Weili Li*
Published : September 30, 2019
Journal of Food & Nutritional Sciences
Elena Castell-Perez*, Rosana G. Moreira, Hal S. Knowles, III
Published : October 01, 2019
Journal of Food & Nutritional Sciences
Nagako Okuda, Makoto Miura, Kazuyoshi Itai, Takuya Morikawa, Junko Sasaki, Tamami Asanuma, Mikako Fujii, Akira Okayama
Published : March 27, 2019
Journal of Food & Nutritional Sciences